
General characteristics of trematodes (Trematoda, or Digenea) |
||
AuthorsEditor |
Trematodes or digeneans is a subclass of obligate endoparasitic flatworms. Digeneans are one of the most widespread and species-rich groups of parasitic invertebrates comprising over 2700 nominal genera and about 18 000 species. Digeneans have a complex life cycle with two or three hosts: first intermediate, second intermediate (supplementary) and definitive. The first intermediate host is a gastropod or bivalve mollusc, or, in rare cases, polychete. The role of the second intermediate host can be played by a variety of animals (fish, amphibians, molluscs, oligochetes, leeches, arthropods, etc.). The definitive host is always a vertebrate. In vertebrates the digeneans are usually localized in the digestive tract and associated glandular organs, but can also invade other organs and tissues including lungs, kidneys, circulatory system, body cavity and reproductive passages. About 12 species of digeneans parasitize humans, of which the most widespread and life-threatening are schistosomes. The disease caused by schistosomes (schistosomiasis) occurs in 74 countries and accounts for about 500 000 deaths each year. |
Contents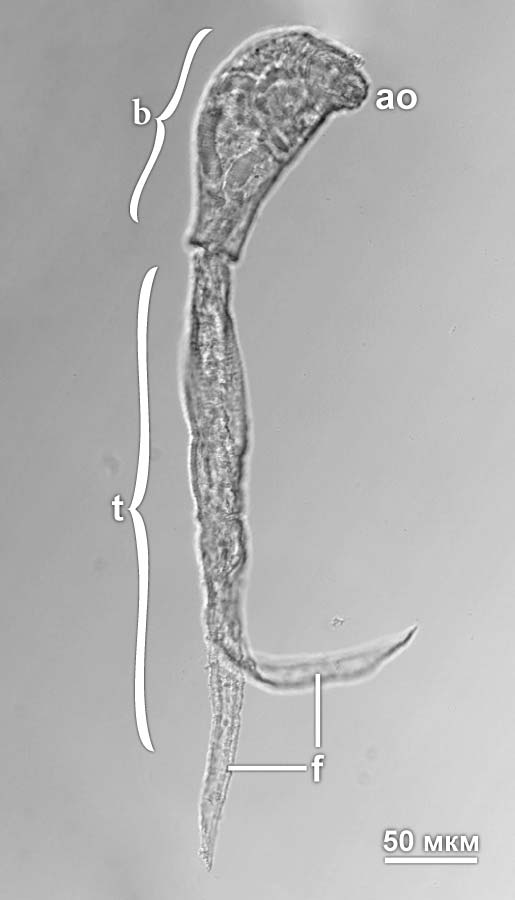
Fig. 1. Cercaria of Diplostomum pseudospathaceum. Abbreviations: ao – anterior organ (glandulo-muscular organ that transforms into the oral sucker in metacercariae); b – body of cercaria; f – furca of the tail; t – tail. Photo by A.A. Petrov

Fig. 2. Generalized morphology of an adult digenean. Abbreviations: ac – ventral sucker (acetabulum); at – common genital atrium; cr – cirrus; cs – cirrus sac; ec – ejaculatory duct; eo – ejaculatory opening; ex – excretory bladder; gb – gut branch; mg – Mehlis’ gland; mo – mouth opening; ph – pharynx; pp – prepharynx; os – oral sucker; ot – ootype; ov – ovary; sr – seminal receptacle; te – testis; vc – vitellary duct; vf – vitellary follicle.
|
Alternation of generations |
Digeneans have complex life cycles that involve the alternation of generations. The life cycle normally includes one hermaphroditic and two parthenogenetic generations. The eggs laid by the adult digeneans are released into the environment, where they hatch into free-swimming ciliated larvae of the first parthenogenetic generation (miracidia). The miracidium penetrates the first intermediate host casting away its ciliated cells and undergoes the metamorphosis transforming into the parthenogenetic stage (primary sporocyst). The sporocyst has a simple morphology and is essentially a sac enclosed in the body wall. The central cavity of this sac houses the embryos that give rise to rediae, the individuals of the second parthenogenetic generation. Rediae are more complex than sporocysts; they have a functional digestive system and can move inside the host’s body. Some digeneans lack this stage, and the primary sporocysts produce instead the second generation of sporocysts (secondary sporocysts). The embryos that grow inside the rediae or secondary sporocysts develop into cercariae, which leave the body of the intermediate host. Cercaria (Fig. 1) is a larva of the hermaphroditic generation that bears a muscular tail at the posterior end; this tail is used for swimming. In most digeneans, the cercariae actively seek their host. Once they have found the host, the cercariae penetrate into its body losing their tails and develop into metacercariae. The second intermediate host is swallowed by the definitive host and metacercariae continue their development in this new host turning into adults. In some groups of digeneans, the second intermediate host is absent and cercariae infect directly the definitive host. In primitive families, the larvae encyst on water plants and remain dormant until the cyst is ingested by the definitive host. |
|
Adult worm |
The body of the adult worm (Fig. 2) usually has a flattened, leaf-like shape. Most worms are less than 5 mm in size, but there are also true giants, like Hirudinella ventricosa that reaches 12 cm in length. Thread-like adult worms of the family Didymozoidae are exceptionally long; the didymozoid Nematobibothrioides histoidii parasitizing the sunfish can grow up to 12 meters in length. The adult worms normally possess two suckers: the oral sucker encircling the mouth and the ventral sucker (acetabulum), but some digeneans may lack one or both suckers. The body of the worm is covered by the syncytial tegument, which is usually beset with spines composed of crystallized actin. The space between the internal organs is filled with parenchyma, which in some digeneans is crossed by a network of canals called the lymphatic system; this system is likely to serve for transportation. The body-wall musculature normally consists of three layers: outer circular, intermediate longitudinal and inner diagonal, but there are different variations in body-wall muscle patterns. The digestive tract is a blind sac and usually consists of a short prepharynx, muscular pharynx, esophagus and two (in rare cases one) branches of the gut. In some digeneans, the gut branches are fused into a single loop. The excretory system is composed of protonephridia; the protonephridia are united into common excretory ducts, which empty into a urinary bladder opening outside through an excretory pore in the posterior portion of the body. In large-bodied species, the epithelium of the bladder can form a network of canals in the parenchyma (paranephridial plexus), which perform the metabolic or excretory function. The reproductive system usually occupies the major portion of the worm’s body and is very complex. The vast majority of trematodes are hermaphrodites; gonochorism is found only in schistosomatids and some didymozoids. The male portion of the reproductive system consists of one or, more often, two testes. The testes give rise to the efferent ducts uniting into a single duct that has a copulatory organ (cirrus) at its distal end. The female portion of the reproductive system comprises an ovary, vitellaria (vitellary follicles) and a complicated system of ducts and glands that participate in the formation of egg envelopes. As a mature oocyte leaves the ovary, it enters the distended portion of the oviduct (ootype), where it becomes fertilized and the egg envelopes begin to form. The ootype is surrounded by Mehlis’ glands, whose secretion serves as a matrix for the components of the egg shell. Mehlis’s glands probably also secrete the lubricant that facilitates the passage of eggs through the reproductive tract. In the ootype, the oocyte becomes surrounded with vitelline cells that reach it via the vitelline duct opening into the ootype. The vitelline cells form the egg shell and provide nourishment for the developing embryo. The formation of the egg continues in the uterus, which opens outside in the anterior portion of the body; the opening of the uterus is often located in the common genital atrium along with the male genital pore. |
|
Sense organs |
The most common and characteristic sense organs of digeneans present in all stages of their life cycle are numerous sensory receptors in the tegument. They are very diverse in structure and include uniciliate sensilla, ciliary pits and nonciliate papillae. Different types of tegumental receptors are likely to have different receptive functions. These sense organs may function as chemo-, mechano-, thermo- or rheoreceptors, but there have been no detailed studies of their specific functional role in digeneans. Tegumental receptors are usually scattered over the whole surface of the body, but they are more densely arranged on the suckers and penetration organs. Their arrangement is species-specific and is widely used in species identification. Most miracidia and many cercariae have two or three inverted eyespots. These eyespots are composed of receptive rhabdomeric cells and melanin-containing pigment cells. Some miracidia and cercariae possess ciliated lamellar organs, which may also function as photoreceptors. |

Fig. 3. Serotoninergic nervous system of a metacercaria of Diplostomum pseudospathaceum stained with antibodies to serotonin. Abbreviations: an – nerves innervating the ventral sucker; cg – cerebral ganglion; de – dorsal nerve cord; on – nerves innervating the ventral sucker; tc – transverse commissures; ve – ventral nerve cord. Photo by A.A. Petrov

Fig. 4. RFamide-ergic nervous system of a metacercaria of Diplostomum pseudospathaceum stained with antibodies to FMRFamide. Abbreviations: cg – cerebral ganglion; de – dorsal nerve cord; le – lateral nerve cord; on – nerves innervating the oral sucker; tc – transverse commissures; ve – ventral nerve cord. Photo by A.A. Petrov
|
Nervous system |
The nervous system of the adult digeneans has an orthogonal architecture typical for the flatworms. The brain is positioned above the pharynx and is composed of a pair of cerebral ganglia connected by the suprapharyngeal commissure. Most species studied to date possess four pairs of anterior nerves; three of those (dorsal, lateral and ventral) arise from the cerebral ganglia and innervate primarily the oral sucker and sense organs at the anterior end of the body, and the fourth pair emerges from the brain commissure and innervates the pharynx. Three longitudinal pairs of nerve cords (ventral, lateral and dorsal) arise posteriorly from the brain. The ventral pair is more developed than the other two pairs; these cords run the length of the worm and in many digeneans connect with each other posteriorly to form a loop. Lateral and dorsal pairs are often shorter than the ventral cords and in some groups can be partially or completely reduced. The nerve cords are connected by transverse commissures arising at regular intervals along the length of the cords. The cords and commissures are associated with peripheral nerve plexuses that innervate the body-wall musculature and internal organs. In some groups of digeneans, the body of the worm is subdivided into two regions: forebody and hindbody; the latter houses the reproductive organs. These worms have an additional ganglion at the posterior end of the forebody that innervates the hindbody. The architecture of the nervous system in cercariae and metacercariae (Fig. 3 and 4) is generally the same as in adults. The cercarial tail usually has a well-differentiated nervous system: in some species the cords continue directly into the tail, in others they end in a nerve plexus at the posterior end of the body; this plexus gives rise to two pairs of nerves that continue into the tail. The nervous system of the miracidia consists of a large, often bilobed, cerebral ganglion and a pair of nerve cords. In sporocysts, the nervous system is simple, but in some digeneans the sporocysts appear to have a cerebral ganglion and one pair of longitudinal nerve cords. The nervous system of rediae is more complex than that of sporocysts; in addition to the cerebral ganglion, rediae can also possess two pairs of anterior nerves and two pairs of nerve cords. The cells of the nervous system of digeneans are mostly bipolar and multipolar. Serotonin (5-HT) appears to be the primary excitatory neurotransmitter of the musculature (Fig. 3), while acetylcholine is the major inhibitory neurotransmitter. This sets parasitic flatworms apart from most other animals, both vertebrates and invertebrates, in which acetylcholine has an excitatory action on muscle cells. The nervous system of digeneans was also shown to contain neuropeptide F and FMRFamide-like peptides (Fig. 4). These peptides are implicated in the control of muscle contraction. |
|
Recommended literature |
|
|