
Pogonophora (Annelida) |
||
AuthorEditor |
Pogonophorans are sedentary marine worms that lack an intestine. They live in tubes made from protein and chitin, which they secrete. Pogonophorans occur in all oceans and many of the marginal seas at depths from 22 to more that 8000 m. They generally prefer cold waters. About 80% of the species occur at depths between 200 and 4000 m. Pogonophorans live primarily on soft silty bottoms, especially in reducing sediments, burrowing deeply (subclass Frenulata). Some prefer to live in decaying organic matter, or sediments rich in methane and sulfide (subclass Monilifera), others in warmer hydrothermal vent conditions or cold seeps (subclass Vestimentifera). There are currently about 170 species of Pogonophora, 90% of which belong to the subclass Frenulata. Pogonophorans get their energy primarily from chemoautotrophic methane- or sulfur-oxidizing bacteria, living endosymbiotically in a specialized organ called the trophosome. They also get some nourishment from dissolved organic matter in the sediment pore water. |
Contents
Representative pogonophorans
General view of the vestimentiferan Riftia pachyptila Jones, 1981. Photo by A.V. Ivanov

Siboglinum scotiense Smirnov, 2015. Tentacles, the forepart and a part of the trunk ventrally. Photo by R.V. Smirnov

Heptabrachia ctenophora Ivanov, 1962. Tentacles, the forepart and a part of the trunk ventrally. Photo by R.V. Smirnov
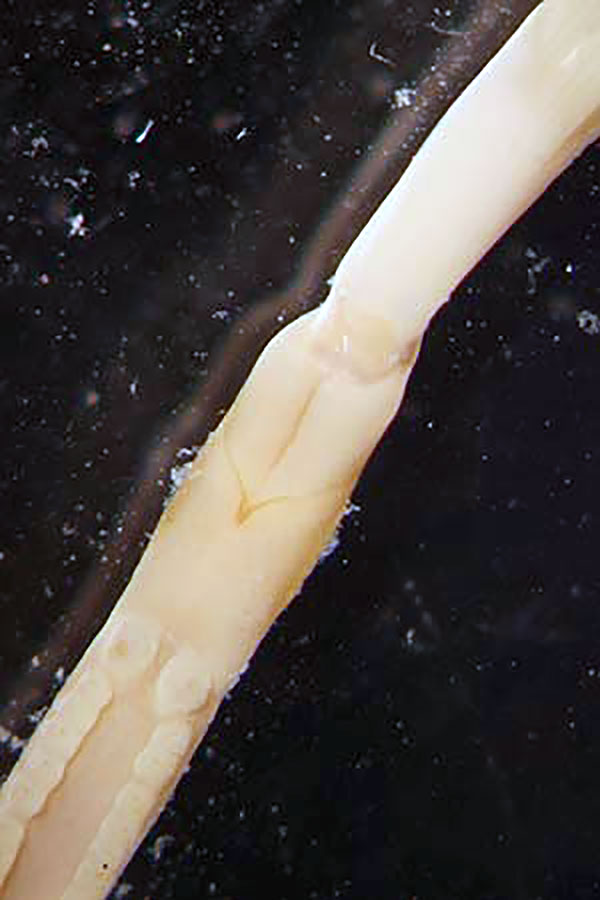
Spirobrachia orkneyensis Smirnov, 2000. Tentacles, the forepart and a part of the trunk ventrally. Photo by R.V. Smirnov
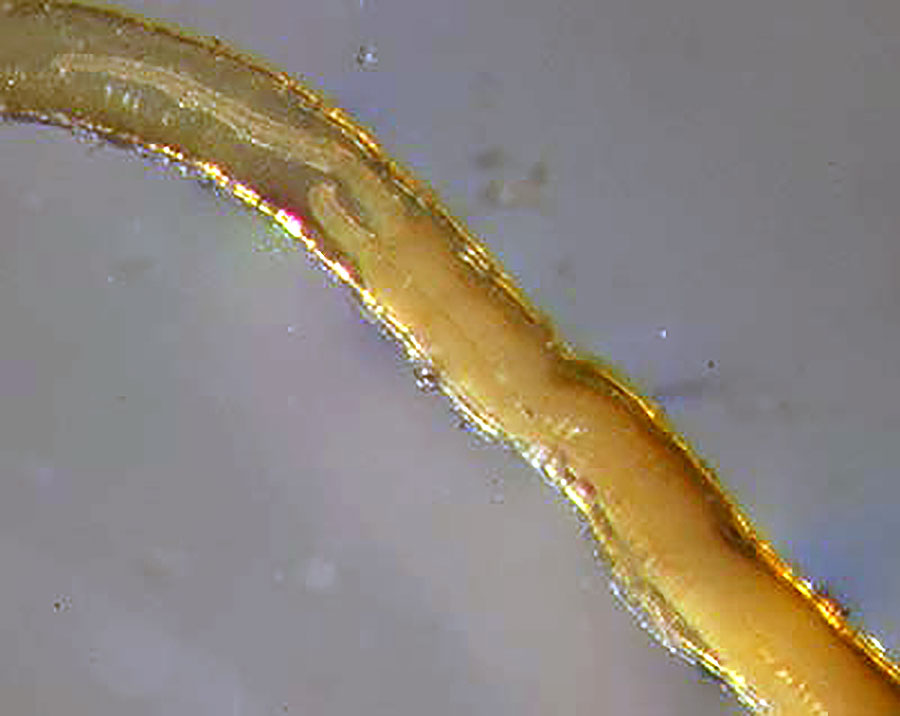
Archeolinum contortum (Smirnov, 2000). Tentacles, the forepart and a part of the trunk ventrally. Photo by R.V. Smirnov
“
The central nervous system of pogonophorans consists of a brain and ventral nerve cord.. Photo by R.V. Smirnov
|
Body segments |
The body is bilaterally symmetrical, remarkably long (the length exceeds the width by hundreds of times), and it has several segments and sections. Pogonophorans are polymerous. The first segment (peristomium) consists of the cephalic lobe (or prostomium) and tentacles (palpi) with a coelomic canal; there can be one to several hundred of such tentacles. The tentacles often have pinnules, which consist of outgrowths of individual epidermal cells (in frenulates) or they are formed of many cells (in vestimentiferans). The bridle (a thickened area of the cuticle found in frenulates), a row of cuticular plates (plaques, found in moniliferans), or vestimental wings (in vestimentiferans) occur on segment two. The coelomic space of this and subsequent segments is filled with numerous multicellular glands. The cephalic lobe and segments one and two make up the anterior section (so-called forepart), which is often demarcated from the rest of the body by a transverse constriction. A large portion of the body consists of segment three, the trunk or metasome. In Frenulata this segment has a complex regional morphology. The anteriormost “metameric” section of the preannular region is characterized by more or less regular paired papillae or dorsal ridges with a groove between them and a ciliated band (the neurotroch) on the ventral side, which present also on the forepart in moniliferans and vestimentiferans. Following this, the evolutionary most ancient nonmetameric part of the preannular region is covered with irregularly distributed papillae. The zone of thickened papillae is a peculiar portion of the posterior preannular nonmetameric region, present in many frenulates. It is characterized by a longitudinal row of regular enlarged papillae on the dorsal side and a ciliated band on the ventral side. The girdles, or annuli, are transverse rows of short bristles (chaetae) developed from the primary set of segmental chaetae, visible in early larvae. The trunk of moniliferans and vestimentiferans is covered with scattered papillae, and is equivalent to the nonmetameric preannular region of frenulates. Girdles are present at the posterior end in moniliferans. A postannular region of the trunk is present only in Frenulata and is characterized by great length and a variety of metameric papillae and glandular regions of the epidermis. The body of all pogonophorans ends with an opisthosoma, consisting of several (to several tens) of segments, and a posterior zone of growth (the pygidium), which can be simple or surrounded by one or two lobes. The segments have a polychaetous standart set of chaetae (two groups dorsolaterally and two ventrolaterally) that are more numerous in moniliferans and vestimentiferans than in frenulates. The chaetae of the opisthosoma and annuli can differ morphologically. Those on the annuli have broad dentate heads in frenulates; the denticles on their heads are arranged in two groups, directed anteriorly and posteriorly. Between these groups there is often a noticeable intermediate zone lacking denticles. On the opisthosoma of frenulates the chaetae are rod- or club-shaped, while in moniliferans and vestimentiferans they are more like frenulate girdle chaetae. |
|
Reproductive system
|
Pogonophorans have separate sexes (with the exception of one species), but external differences between males and females are not obvious in frenulates or moniliferans. Some frenulates have anterior enlarged genital papillae, and vestimentiferans have external genital grooves, showing differences between males and females. The sperm are packed into true spermatophores (subclass Frenulata) or sperm masses (spermatozeugmata) (subclass Vestimentifera, some species) or are free (subclasses Monilifera and some Vestimentifera). The spermatophores often have a variety of processes (wings, threads, etc.). The oocytes (eggs) are exuded into the space between the body and the tube wall, and are transported by means of the ciliary band to the anterior of the tube where they are either released into the water or undergo initial stages of embryonic development to the point of forming trochophore-like lecithotrophic larvae or even juveniles. Fertilization may be internal or external. The release of small eggs is characteristic of vestimentiferans and moniliferans, and also the order Thecanephria of Frenulata. Brooding of embryos in the tube is known in some members of the order Athecanephria of Frenulata. Planktotrophic trochophores and crawling, feeding larval (juvenile) stages are known in Vestimentifera. Larvae are not known in the Monilifera and Thecanephria. Pogonophorans with different development types have different reproductive strategies with corresponding features of the female gametes. Large, elongate, yolky eggs in low numbers are characteristic of some Athecanephria; for all other groups of Pogonophora, the eggs are small, round, numerous, and less yolky. |
|
Excretory
|
Pogonophorans have a protonephridium-type excretory system. The excretory pores are located at the anterior part of the body on the dorsal or ventrolateral sides. The circulatory system is closed with a dorsal heart, sometimes enclosed in a coelomic sac (a pericardium); a cardiac body is often present in the medial blood vessels. |
|
Nervous system |
The completely intraepidermal nervous system is situated in the cutaneous epithelium. The central nervous system is represented by a poorly morphologically differentiated brain and a longitudinal unpaired nervous tract on the ventral side of the body. The brain is simply a broadening of the trunk ventral nervous tract combined with a concentration of nervous elements in the cephalic lobe and in the first segment encircling the body like a larval peripharyngeal (prototrochal) nervous ring. Initially the brain is medial-apical in larvae and early juveniles of Pogonophora. Some early vestimentiferan juveniles possessing a functioning intestine, have the brain penetrated by the anterior gut. Since the ventral or dorsal position of the brain could be determined only in relation to the pharynx and the anterior gut, the absence of the latter and the small size of majority of pogonophorans make the brain possible to be displaced closer to the ventral nervous tract to take the medial-ventral position. Though the nervous tract is morphologically unpaired, immunohistochemical experiments and embryological studies revealed its initially paired nature throughout the whole body of the adult worm. In addition, the nervous trunk in the opisthosoma is ladder-like, with three longitudinal tracts and ganglia in each segment, while vestimentiferans have the nervous trunk bifurcated in the area of the vestimentum. These peculiarities of the pogonophoran nervous system do not fit the Polychaeta classic neuromerous scheme, but agree well with the structure of the nervous system in many groups of so-called Archiannelida. Both pogonophores and archiannelids have simple nervous system consisting of intra- or subepithelial general plexus and paired (or unpaired) tract without ganglion-like concentrations of nerve cells. Pogonophorans are more elaborated in this respect, because their larvae and adults (in opisthosoma) have ganglia. In pogonophoran family Spirobrachiidae and in archiannelid family Polygordiidae the nervous tract shows a tendency to submerge into the epidermis in a sulcus, while vestimentiferans demonstrate the initial submersion of the nervous system by the growing of the epidermal layer into the connective tissue, that is characteristic of the majority of polychaetes. Pogonophoran larvae and adults have photoreceptors of the phaosome type, else found in hirudineans and oligochaetes. |
|
Specialization
|
The pinnules, separate cells in the epithelium of the tentacles in Frenulata with internal blood capillaries, are unique structures demonstrating extremely deep specialization of the pogonophoran circulatory system for respiration and transport of some matters (methane, hydrogen sulfide) dissolved in water to endosymbiothic chemoautotrophic bacteria. The absence of the gut in pogonophoran adults (and in early embryonic stages in some species) is a striking example of specialization of nonparasitic animals. The body plan of Pogonophora is peculiar also because of an original multilevel regionation of the body, which firstly originates on the base of elongation of some segments (but not of fusion of segments like in many sedentary polychaetes), following by the irregular growth of different parts of the trunk accompanied with metamerism of different ectodermal organs. In addition, the heart and the chitin-protein tube are also synapomorphic for Pogonophora. |
“
The lack of the gut and the presence of pinnules (cells in the tentacular epithelim with capillaries inside) are features showing a high degree of specialization of pogonophorans.
|
Tube |
The tubes of Frenulata are nearly cylindrical and straight or curved and are normally not branched; they are rarely closed at the anterior or posterior ends and often have some kind of ornamentation. Most often one observes ringed tubes, more rarely ringed-segmented or segmented tubes. The rings are the areas composed of the densest and darkest colored material; the segments are the result of episodic growth and are limited by narrow encircling constrictions, wrinkles, or overlaps, sometimes with funnel-like edges, repeated at even intervals. The anterior ends of the tubes are often soft and filmy, sometimes in the form of rigid funnels. The tubes of Monilifera lack rings or segments; their shape is often very convoluted and their surface finely wrinkled. The tubes of vestimentiferans are segmented and funneled anteriorly; they are non-ringed, of tough construction, and usually closed at the very narrow posterior end, at least in the early stages of growth. |
|
References |
|
|
