Тип Rotifera – Коловратки |
||
АвторыРедактор |
Коловратки – разнообразная группа мелких псевдоцеломических животных, повсеместно распространенных в пресных водоемах и в морях. Встречаются они и среди сфагнума в болотистой местности и на мхе во влажных лесах. Известно около 2000 видов коловраток (Segers, 2007). Большинство свободноподвижные, но есть и прикрепленные формы. Небольшую группу коловраток составляют виды, использующие в качестве субстрата других животных и растения. Среди коловраток лишь несколько десятков представителей достигают в длину 1–2 мм, остальные существенно мельче. В среднем их длина 0.1–0.5 мм. Колонии коловраток могут достигать 3-4 мм в диаметре. Коловратки обладают высокоспециализированными тканями и постоянством клеточного состава, иначе эутелией. Характерной особенностью коловраток является наличие особого коловращательного аппарата, получившего название короны. Он представляет собой снабженный собственной мускулатурой ресничный орган, используемый животным для захвата пищи и передвижения. Биение ресничек короны напоминают мелькание спиц крутящегося колеса, отсюда и латинское название коловраток – Rotifera – колесонесущие. Туловище животных содержит внутренние органы, в том числе и склеротизированный челюстной аппарат или иначе мастакс. Тело коловраток имеет мускулистый вырост - ногу, благодаря которой животные могут ползать или прикрепляться к субстрату. На конце ноги часто располагается пара выростов – «пальцев». Коловратки бывают как беспанцирными (рис. 1A), так и панцирными (рис. 1B). По способам движения они разделяются на 4 группы: плавающе-ползающие, плавающие, ползающие и прикрепленные к субстрату. |
Содержание
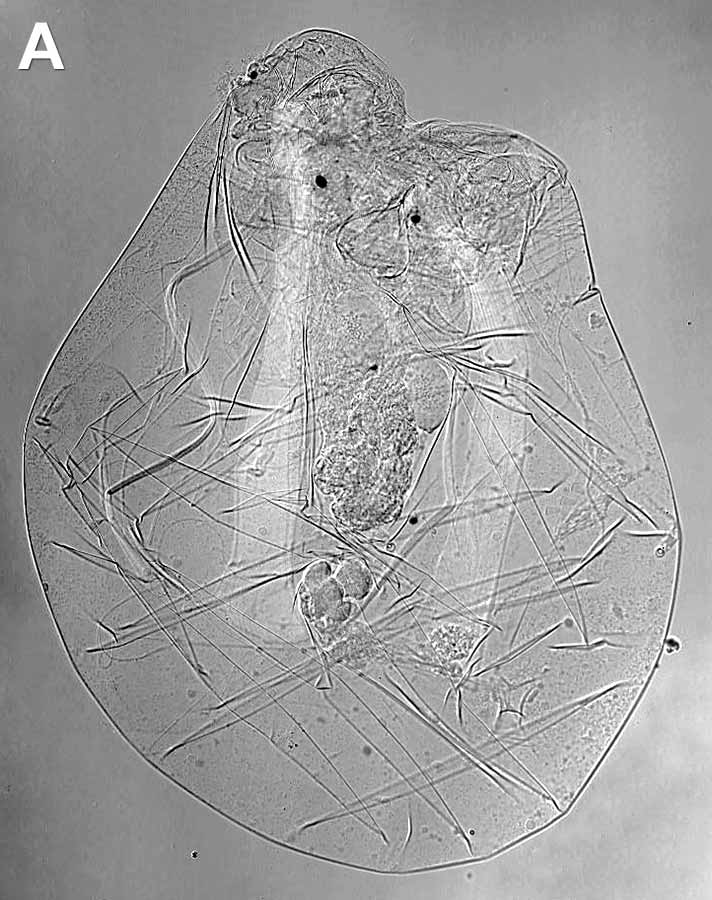

Рис. 1. Общий вид коловраток: А – беспанцирная Asplanchna priodonta, B – панцирная Brachionus sp. Метод дифференциально интерференционного контраста по Номарскому (DIC). Фото Е.А. Котиковой

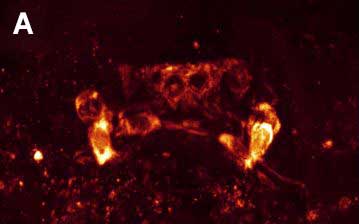
Рис. 3. Двойное ICC окрашивание нервной системы коловратки Platyias patulus. FMRFамид-IR показана красным, а 5-HT-IR показана зеленым. Фото Е.А. Котиковой
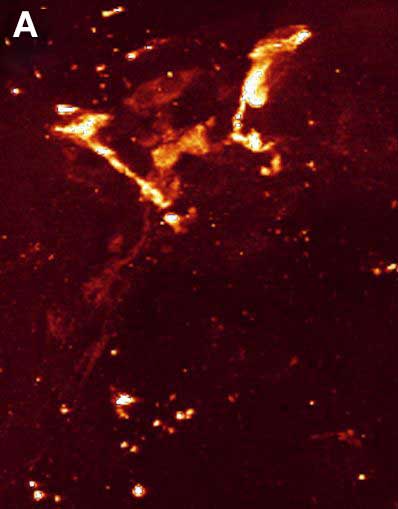


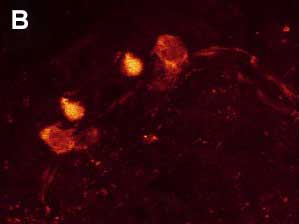
Рис. 4. Двойное ICC окрашивание нервной системы коловратки Euchlanis dilatata. A – FMRFамид-IR. B – FMRFамид-IR показана красным, а 5-HT-IR показана зеленым. C – 5-HT-IR. Фото Е.А. Котиковой
|
Классификация коловраток |
Коловратки раздельнополые животные. Половая система самок представлена цельным (Monogononta) или двулопастным (Digononta) яичником. Именно этот факт послужил в конце 19 века разделению всех коловраток на две соответствующие группы (Wesenberg-Lund, 1899). Эта классификация коловраток сохранилась до настоящего времени. Русскими исследователями была предпринята попытка создания другой системы, основанной на морфофункциональном подходе к изучению ключевых структур коловраток: короны и мастакса (Кутикова, 1970; Маркевич, 1991). К сожалению, новая система не нашла своих последователей на Западе и остается малоизвестной. В последнее время приоритет получили молекулярно-филогенетические исследования, основательно пересмотревшие филогению билатерально симметричных животных (Bilateria), но и они не прояснили положение коловраток полностью. Для Rotifera вместе с Acanthocephala (скребни) предложено название Syndermata, которое подчеркивает их синапоморфию, а именно, уникальный синцитиальный эпидермис с лежащей в цитоплазме пластинкой (Ahlrichs, 1997; Barnes et al., 2001). Однако, до сих пор филогенетические отношения внутри Syndermata остаются не уточненными. В недавнем исследовании Фонтането и Йонделиуса (Fontaneto, Jondelius, 2011) изучены более тысячи последовательностей COI и поддержаны клады Acanthocephala и коловраток: Bdelloidea, Monogononta и Seisonidea. Взаимоотношения этих групп остались неясными, за исключением четкого выделения сестринских отношений между Acanthocephala и Bdelloidea. |
|
Нервная система |
Первым завершенным этапом в изучении всех систем органов коловраток, включая нервную систему, стали классические гистологические исследования, результаты которых были собраны и проанализированы А. Ремане (Remane, 1929-32). Позднее изучение ультраструктуры коловраток с привлечением электронной микроскопии выявило ряд новых особенностей организации их нервной системы. В нейронах мозга коловраток было описано по крайней мере 4 типа синаптических пузырьков (Villeneuve, Clément, 1971; Clément, 1977; Wurdak et al., 1983). Самые мелкие из них имели электронно-прозрачную сердцевину и могли содержать ацетилхолин, наличие которого было подтверждено позднее биохимическими и гистохимическими исследованиями без картирования нейронов (Nogrady, Alai, 1983). В других нейронах мозга были выделены пузырьки средних размеров с электронно-плотной сердцевиной, которые могли содержать катехоламины (КА) или серотонин. Комплекс трех различных био- и гистохимических методик выявил локализацию КА в нервной системе коловраток, но опять не позволил показать расположение этих нейронов в нервной системе (Keshmirian, Nogrady, 1987, 1988). В начале 90-х годов ХХ века в лаборатории эволюционной морфологии Зоологического института РАН были начаты работы по исследованию распределения КА в нервной системе коловраток. Основной методикой этих исследований был выбран гистохимический водно-глиоксиловый метод (GIF) в модификации Каботянского (1985) с пятикратным увеличением концентрации кислоты в инкубационной среде. Объектами исследования послужили 12 видов коловраток, из которых 2 из одного семейства в группе Digononta и 10 из 7 семейств Monogononta, далеко отстоящих друг от друга (Kotikova, 1995; 1998a, b). Основные вариации в строении КА-ергической нервной системы у коловраток наблюдаются в расположении нейронов в мозге (рис. 2). 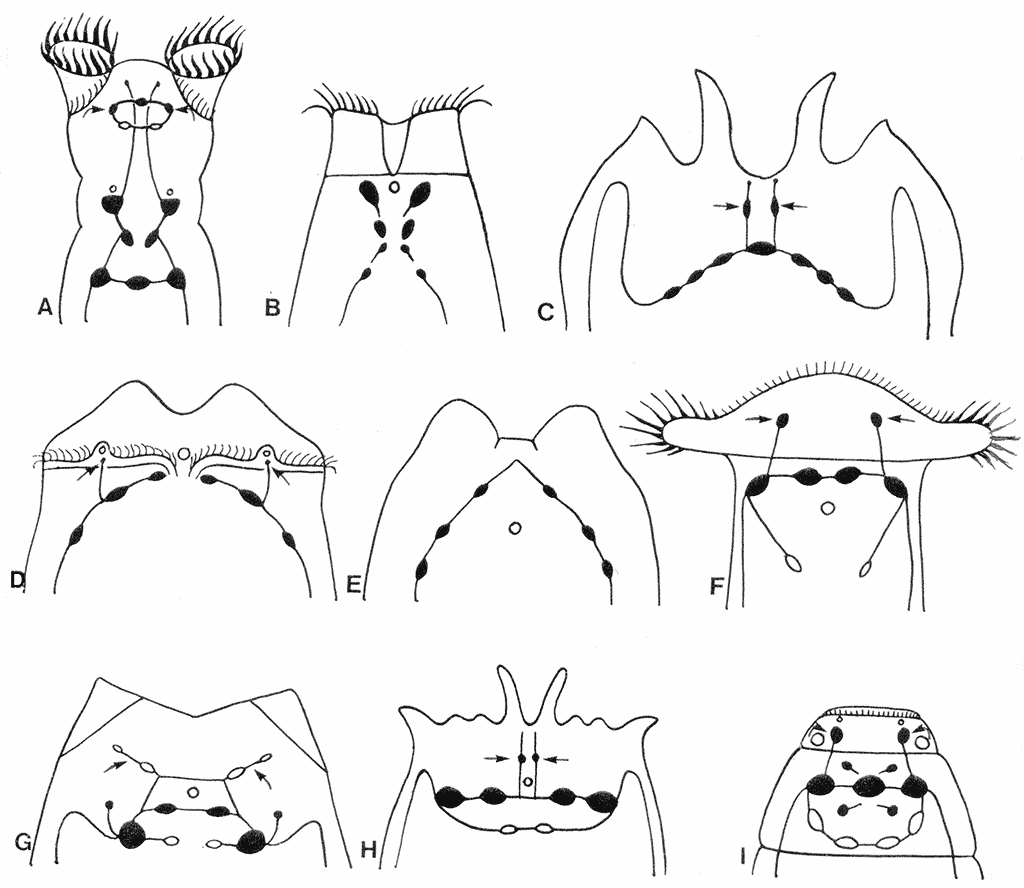
Рис. 2. Схема расположения КА-ергических нейронов (черные) в мозге коловраток. A – Philodina sp. B – Manfredium eudactylotum. C – Platyias quadricornis. D – Asplanchna herricki. E – Euchlanis dilatata. F – Notommata sp. G – Lecane arquata. H – Brachionus quadridentatus. I – Dicranophorus forcipatus
Выделено три ступени усложнения в геометрии расположения мозговых нейронов: Х-образная (трех или четырех ярусная), дугообразная (плавная или крутая) и кольцеобразная. Мозговые нейроны у Philodina и Rotaria (рис. 2А) могут вытягиваться в продольном направлении в три или четыре яруса, образуя Х-образные структуры (Kotikova, 1995). У коловраток с дугообразным мозгом плавный изгиб отмечен у Platyias (рис. 2С), а более крутой у Euchlanis (рис. 2E). Переход от дугообразной к кольцеобразной форме мозга осуществляется в два этапа. На первом отмечается незначительная степень закругления, выраженная в образовании полукольца у Lecane (рис. 2G). Второй этап приводит к появлению замкнутого эллипсовидного мозгового комплекса у Brachionus и Dicranophorus (рис. 2H, I). Во всех случаях от боковых нейронов мозга начинаются вентролатеральные стволы. Кроме нейронов в мозге отмечено наличие нейронов по ходу стволов, а также в области головы и мастакса (глотка коловраток с особым жевательным аппаратом). Нейроны мозга коловраток составляют 20% от общего числа всех их клеток (Martini, 1912). Применение компьютерной программы трехмерной реконструкции продемонстрировало четкую билатеральную симметрию расположения 200 нейронов мозга у Asplanchna brightwelli (Ware, Lopresti, 1975). Общее число КА-ергических мозговых нейронов в нашем материале незначительно (6-11), но постоянно для каждого вида и составляет 3-6% от общего количества нейронов мозгового комплекса. И только у Notommata copeus ICC выявлено 40 перикарионов в мозге, содержащих FMRFамид и 5-HT IR (Hochberg, 2007), а у Notommata sp. четыре КА-ергических мозговых нейрона (Kotikova, 1998a), что в совокупности составляет уже 22%. Наличие не уточненных по химическому составу синаптических пузырьков послужило стимулом для начала иммуноцитохимических исследований нервной системы коловраток, для выявления других нейротрансмиттеров. |
|
Иммуноцитохими-ческое исследование нервной системы |
Объектами иммуноцитохимического (ICC) исследования c применением антител к полипептиду FMRFамиду и моноамину серотонину (5-HT), послужили три вида коловраток из высокоорганизованного отряда Ploima. Это плавающее-ползающие панцирные Platyias patulus, Euchlanis dilatata и беспанцирная плавающая Asplanchna herricki. У Platyias patulus (рис. 3) восемь мозговых FMRFамид IR нейронов расположено дугообразно в 2 яруса: пара мелких биполяров находятся на переднем дорсальном полукольце, пара униполяров - в области короны и 2 пары нейронов - по ходу вентро-латеральных продольных стволов (Kotikova et al. 2005). В мозге выявлено шесть 5-HT IR нейронов, расположенных Х-образно в три яруса, 2 пары биполяров на переднем дорсальном полукольце и пара биполяров по ходу продольных стволов (рис. 3). Волокна продольных стволов FMRFамид- IR и 5-HT IR следуют параллельно друг другу, а нейроны с той же IR располагаются независимо друг от друга в мозге и по ходу стволов. У Euchlanis dilatata 6 пар FMRFамид IR нейронов (Kotikova et al., 2005): две пары мозговых нейронов лежат дугообразно; по ходу стволов выявлено еще 2 пары нейронов, а в области короны – пара мультиполяров (рис. 4). В боковых участках мозга 2 пары 5-HT-IR нейронов лежат дугообразно, по ходу стволов выявлено 2 пары биполярных нейронов. У Asplanchna herricki (Kotikova et al., 2005) четырнадцать мозговых FMRFамид IR нейронов формируют кольцеобразную структуру: 6 имеют форму шестиугольника (вентральная сторона мозга), а 8 лежат на дорсальной стороне и имеют каплевидную и сферическую форму. По ходу стволов лежит еще пара биполярных нейронов (рис. 5А). В дугообразном нейропиле мозга присутствует также две пары 5-HT IR нейронов: более мелкие срединные (3.5 мкм) имеют яркую окраску, тогда как более крупные (5 мкм) сдвинуты в латеральном направлении и окрашены слабее. Вентролатеральные стволы включают две пары нейронов: проксимальные и дистальные (рис. 5B). У всех исследованных видов двойное ICC окрашивание продемонстрировало независимое расположение мозговых нейронов размерами от 2 до 10 мкм. Волокна различной ергидности всегда следуют параллельно друг другу, причем FMRFамид IR проходят под 5-HT IR волокнами. Ко-локализации нейротрансмиттеров не выявлено. Через год после публикации наших материалов появилась статья, в которой было описано распределение серотонинергических элементов в нервной системе 2 планктонных коловраток Conochilus coenobasis и C. dossuarius (Hochberg, 2006). У этих видов 5-HT-ергическая нервная система представлена дорсальным мозговым ганглием и 4 парами формирующих кластеры перикарионов, а также парой продольных нервных стволов, соединяющихся в области ноги. Годом позже выходит вторая статья этого же автора, в которой подробно описаны FMRFамид- и 5-HT IR участки нервной системы бентической коловратки Notommata copeus (Hochberg, 2007). Исследуемые медиаторы обнаружены в мозге и паре продольных стволов. Мозговой ганглий включает 30 FMRFамид- и 10 5-HT IR перикарионов. Пептидергические перикарионы располагаются по периферии мозга, демонстрируя слабую окраску, тогда как часть центральных окрашена ярче. Распределение 5-HT IR элементов описано и у бделлоиды Macrotrachela quadricornifera (Leasi et al., 2009). Появление сходной геометрии расположения мозговых КА-ергических, FMRFамид IR и 5-HT IR нейронов у представителей трех разных семейств одного отряда Ploima говорит о независимом и параллельном развитии этих структур у далеко отстоящих друг от друга коловраток. У низших Bdelloida описан один, Х-образный, тип расположения мозговых структур, а у более высокоорганизованных коловраток обнаруживаются все три, а именно Х-образное (2-3-х ярусное), дугообразное и кольцеобразное. |
|
Рекомендуемая литература |
|
|