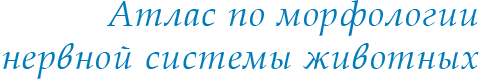
Рептилии (Reptilia) |
Рептилии (Пресмыкающиеся) являются ключевым классом высших позвоночных животных, от которых эволюция пошла по пути формирования двух других классов высших позвоночных – птиц и млекопитающих. Рептилии – первый класс позвоночных, полностью вышедших на сушу (водные черепахи, морские змеи – вторично освоили водную среду), что привело к кардинальным изменениям во всех системах органов животных, включая нервную систему и органы чувств. С позиции эволюционной нейроморфологии рептилии представляют исключительный интерес, поскольку именно в этом классе позвоночных закладываются те особенности организации головного мозга, которые дали начало развитию высших центров мозга птиц и млекопитающих. К этим особенностям относятся:
|
Содержание |
Общий обзор строения нервной системы Рептилий |
||
Спинной мозг |
У рептилий в спинном мозге происходит дальнейшая дифференциация серого и белого вещества. Особенно это касается распределения групп мотонейронов в передних рогах серого вещества. Мотонейроны спинного мозга рептилий организованы в две моторные группы: вентромедиальную и вентролатеральную, нейроны которых отличаются друг от друга по размеру и характеру ветвления дендритов. У рептилий впервые среди позвоночных дендриты большинства мотонейронов не проникают на противоположную сторону спинного мозга, и это позволяет осуществлять более тонкую регуляцию деятельности мотонейронов спинномозгового сегмента через систему интернейронов (рис. 2). Мотонейроны первой группы иннервируют осевую мускулатуру, второй – мускулатуру конечностей. У рептилий мотонейроны, иннервирующие отдельные мышцы, уже расположены раздельно, тогда как у амфибий мотонейронные пулы разных мышечных групп перемешаны. Кроме того, у рептилий (особенно у змей) самая сложная среди позвоночных мышечная система, в связи с чем, у них встречается несколько вариантов моторной иннервации. Есть мышцы, которые иннервируются мотонейронами, расположенными в разных сегментах спинного мозга. Такая иннервация называется полинейронной, причем нейромышечные контакты от разных мотонейронов располагаются по всей длине мышечного волокна. Другие мышцы иннервируются мононейронально и локально. Очень часто в отдельных мышцах красные и белые волокна перемешаны друг с другом и иннервируются разными мотонейронами. Считается, что такая иннервация мышц у рептилий обеспечивает синхронизацию их работы и более тонкую регуляцию (например, при движении туловища у змей). Все проводящие пути спинного мозга сосредоточены в белом веществе, разделяемом на три канатика: задний, боковой и передний. Основной объем канатиков занимают супраспинальные тракты, осуществляющие двустороннюю связь спинного и головного мозга (спинно-ретикулярные, спинно-тектальные, спинно-таламические, спинно-мозжечковые и др.). В отличие от низших позвоночных, у рептилий намечается четкая дифференциация проводящих пучков. Впервые выделяется зона проприоспинальных трактов. Нисходящие супраспинальные тракты (ретикуло-, вестибуло-, текто-, руброспинальные и др.) опосредуют влияние головного мозга на моторные центры спинного мозга и имеют сходное строение у всех позвоночных. |
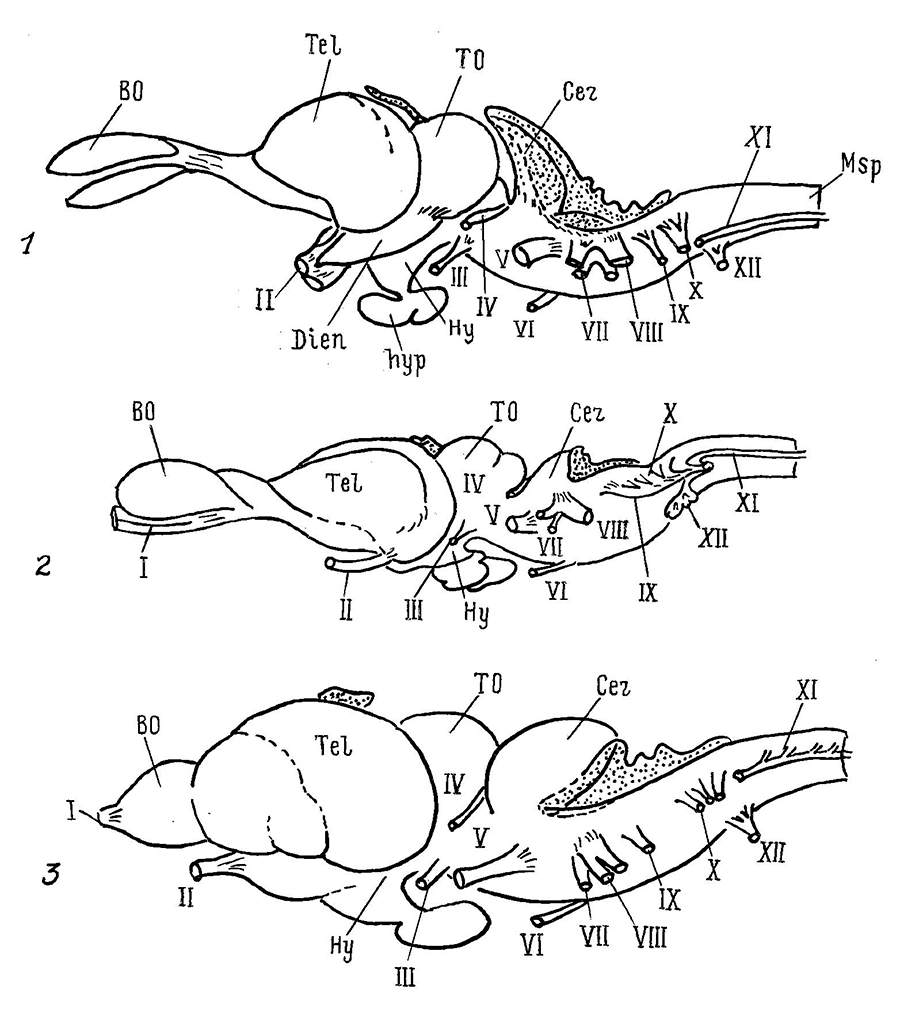
Рис. 1. Общий вид головного мозга некоторых представителей современных рептилий (по: Обухов, Андреева, 2017; Bangma, 1983).
1 – ящерица Varanus exanthematicus, 2 – питон Piton regius, 3 – черепаха Pseudemis ccripta elegans. ВО – обонятельная луковица, Tel – конечный мозг, TO – крыша среднего мозга, Dien – промежуточный мозг, Hy - гипоталамус, hyp – гипофиз, Cer – мозжечок, Msp – спинной мозг, I–XII – черепно-мозговые нервы. 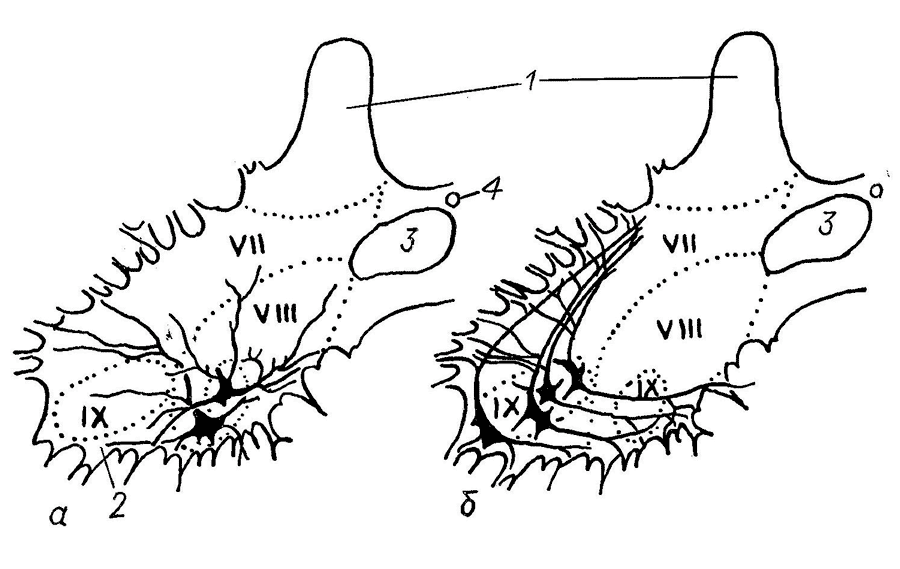
Рис. 2. Нейронная структура вентромедиальной (а) и вентролатеральной (б) группы мотонейронов в спинном мозге ящерицы (по: Cruce, 1979).
1 и 2 – задние и передние, соответственно, рога серого вещества спинного мозга; 3 – медиальные проприоспинальные тракты; 4 – спинномозговой канал; VII–IX – пластины Рекседа |
Продолговатый мозг |
Спинной мозг (Medulla spinalis) имеет сходное строение со всеми позвоночными. Основным принципом организации спинного мозга является его сегментарное строение и внутреннее разделение на серое и белое вещество. Он располагается в позвоночном канале от первого шейного позвонка до крестца, где переходит в краевую нить. По ходу спинного мозга имеется два утолщения: шейное, сегменты которого управляют крыльями и пояснично-крестцовое, связанное с иннервацией задних конечностей. Разделение серого вещества на передние (вентральные) и задние (дорсальные) рога у птиц выражено слабо. Боковые рога практически отсутствуют, в связи с чем, нейроны, связанные с вегетативной нервной системой, располагаются ближе к центральному району серого вещества (рис. 1). Формирующиеся у рептилий и других высших позвоночных связи сенсорных ядер продолговатого мозга с моторными и вегетативными центрами головного мозга обеспечивают такие сложные рефлекторные реакции как жевание, глотание, слюноотделение, рвотный рефлекс. Восходящие связи сенсорных областей мозга связаны с формированием пищевого и питьевого поведения. Наибольшие изменения в продолговатом мозге рептилий связаны с октаво-латеральной системой. Переход к наземному образу жизни привел к редукции центров октаво-латеральной системы, связанных с боковой линией и электрорецепцией и последующим развитием вестибулярных и слуховых центров (VIII – слухо-статический нерв). Формируется сложный комплекс вестибулярных и слуховых ядер, связанных трактами со спинным и средним мозгом. Вентральные отделы продолговатого мозга занимает висцеромоторная зона, которая включает моторные ядра тройничного, лицевого, языкоглоточного и блуждающего нервов. Они обеспечивают иннервацию дыхательной и языко-глоточной мускулатуры. Соматомоторная зона связана с моторными центрами III, IV, VI (глазодвигательные ядра) и XII (подъязычный нерв) черепно-мозговых нервов. Интересно, что среди рептилий наиболее сложным строением моторного ядра XII нерва обладают хамелеоны, отличающиеся наиболее сложным мышечным аппаратом языка. Таким образом, у рептилий, при сохранении общего плана строения продолговатого мозга, наблюдается четкая эволюционная тенденция к усложнению и дифференциации как морфологической структуры, так и системы проводящих трактов мозга. |
|
Задний мозг (Metencephalon) и мозжечок (Cerebellum) |
У рептилий в мозжечке выделяют традиционные для всех позвоночных отделы: латеральные области и медиальную часть. Латеральные области, хорошо развитые у низших позвоночных и связанных с октаво-латеральной системой и получившие наименование ушки мозжечка (auriculae cerebelli) у рептилий практически редуцируются и превращаются в клочково-узелковую долю (lobus flocculo-nodularis). Медиальная часть составляет тело мозжечка (corpus cerebelli), разделяемый бороздами на доли и дольки (рис. 3). В глубине тела мозжечка у рептилий формируются два ядра, нейроны которых образуют систему эфферентных связей. Гистологическая структура мозжечка рептилий типична для всех позвоночных: в коре выделяют молекулярный, ганглионарный (слой клеток Пуркинье) и гранулярный слои. Следует отметить, что формирование четкой слоистой структуры коры в мозжечке характерно именно начиная с рептилий, поскольку у амфибий клетки располагаются по всей толще стенки мозжечка. Клетки Пуркинье мозжечка рептилий имеют крупное грушевидное тело (25–35 мкм) и хорошо развитую систему дендритных ветвлений. Клетки Пуркинье контактируют с системой афферентных лиановидных, моховидных и мультислоистых волокон, связывающих мозжечок со спинным мозгом, структурами ствола мозга и другими центрами головного мозга. Также у рептилий начинает формироваться сложная внутренняя межнейронная структура (например, образование гломерулярных структур в гранулярном слое; появление корзинчатых клеток в молекулярном слое). Эфференты мозжечка образованы аксонами клеток мозжечковых ядер и проходят в ножках мозжечка. По ним информация поступает к ядрам вестибулярного комплекса ствола мозга, красному ядру, нейронам крыши среднего мозга и вышележащим центрам промежуточного (таламус) и конечного мозга. Общим для всех эфферентных трактов мозжечка является то, что через них оказывается влияние мозжечка на моторные центры ствола мозга и спинного мозга. |
|
Средний мозг (Mesencephalon) |
В среднем мозге рептилий выделяют крышу среднего мозга (tectum mesencephali, tectum opticum), покрышку (tegmentum) и ножки мозга (pedunculi cerebri). Полостью среднего мозга является так называемый водопровод (aqueductus cerebri). Тектум изучен у рептилий достаточно хорошо и состоит из системы клеточных и волоконных слоев, число которых варьирует в зависимости от вида животных и степени развития зрительной системы. Наиболее сложное строение тектума встречается у хамелеонов, варанов, агам, ряда видов змей (рис. 4). Основной объем информации тектум получает из сетчатки, причем проекции четко организованы по ретино-топическому принципу: определенные зоны сетчатки проецируются в определенные участки тектума. Развиты связи и с другими отделами головного мозга. Эфференты тектума направляются в вышележащие отделы мозга (в основном в дорсальный таламус), а нисходящие связи направлены в ретикулярную формацию ствола и моторные центры спинного мозга. Последние обеспечивают такие реакции как поворот в сторону добычи/хищника, сочетанные повороты глаз и головы и др. Помимо центров, связанных со зрительным анализатором, у рептилий под тектумом располагается крупная группа нейронов – торус (torus semicircularis), являющийся слуховым центром и гомологом нижних бугорков четверохолмия мозга млекопитающих. В тегментуме среднего мозга располагаются моторные ядра III (глазодвигательного) и IV (блокового) черепно-мозговых нервов, а также крупный центр экстрапирамидной системы мозга – красное ядро (nucleus ruber). Нейроны красного ядра формируют мощные рубро-спинальные и рубро-бульбарные тракты. На границе тегментума и ножек мозга у рептилий находится интерколликулярное ядро, являющееся возможным гомологом черной субстанции (substantia nigra) млекопитающих. Таким образом, средний мозг рептилий содержит нервные центры, связанные со зрительной и слуховой сенсорными системами, а также моторными и премоторными структурами мозга. Все они, в целом, определяют роль среднего мозга, как центра организации и контроля сложных поведенческих реакций рептилий. |
|
Промежуточный мозг (Diencephalon) |
Таламус рептилий, как важнейший центр переключения информационных потоков от сенсорных систем в структуры конечного мозга получает значительное развитие. Это выражается в возрастании количества ядерных образований и четко выраженной направленности проекций таламуса в полушария. Основную трудность представляет задача сравнения структур промежуточного мозга рептилий, птиц и млекопитающих и установления их гомологии. Гипоталамус и эпиталамус во многих отношениях оказались структурами более консервативными, сохраняющие сходные черты организации у всех позвоночных. Это, во-первых, связь с эндокринной системой, реализуемые через нейросекреторные центры гипоталамуса и гипофиза. Во-вторых, конвергенция разнообразной информации и участие гипоталамуса в организации поведенческих реакций. В-третьих, связь гипоталамуса со зрительной системой и участие в регуляции биологических ритмов организма. |
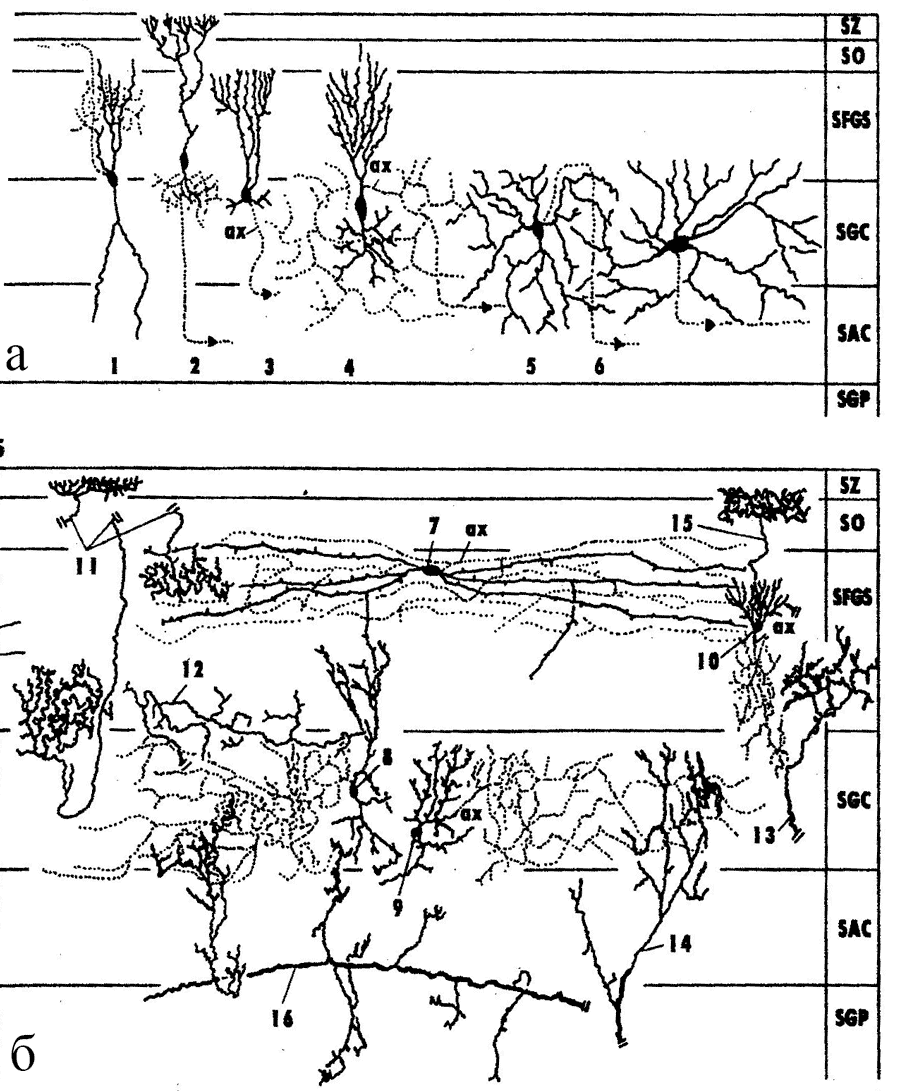
Рис. 4. Структура тектума среднего мозга змеи Thannophis sirtalis (по: Обухов, Андреева, 2017).
а – эфферентные нейроны тектума, б – интернейроны и эфферентные волокна, 1–6 – эфферентные нейроны: текто-геникулятный (1), текто-истмальный (2), текто-истмо-бульбарный (3), текто-таламический (4), текто-бульбарный прямой (5) и перекрещенный (6), 7–10 – различные типы интернейронов тектума, 11–16 – афферентные волокна из сетчатки (11), тегментума (12), латерального коленчатого тела метаталамуса (13), таламуса (14), перешейка (istmus) (15), моторных ядер среднего мозга (16). SZ-SGP – слои тектума: SZ – stratum zonales, SO – stratum opticum, SPGS – stratum fibrosum et griseum superficiale, SGC – stratum griseum centralis, SAC – stratum album centralis, SGP – stratum griseum periventriculare |
Конечный мозг (Telencephalon) |
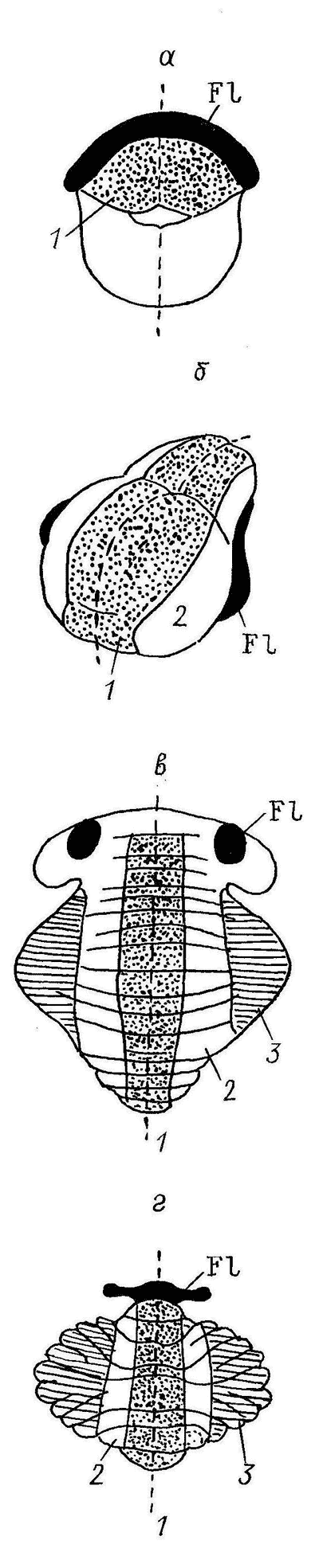
Конечный мозг (Telencephalon) – важнейший отдел ЦНС рептилий. Именно у них формируются формации паллиума, гомологичные корковым структурам мозга птиц и млекопитающих. Паллиум рептилий разделяется на медиальный (архикортекс), дорсальный (возможный гомолог неокортекса) и латеральный (палеокортекс) отделы (рис. 5). Долгое время дорсальный паллиум рассматривали как гомолог неокортекса (слоистая структура, сложные формы нейронов, определенная система связей), однако окончательно этот вопрос не решен. На эту роль претендует и другой отдел полушария: дорсо-вентрикулярный край (ДВК), имеющий паллиальное происхождение, прямые связи с сенсорными ядрами таламуса, сложный нейронный состав. Ряд автором рассматривает ДВК как специфический отдел полушарий мозга рептилий, не имеющий аналога в мозге млекопитающих и развивающийся независимо и параллельно с другими кортикальными формациями других высших позвоночных. 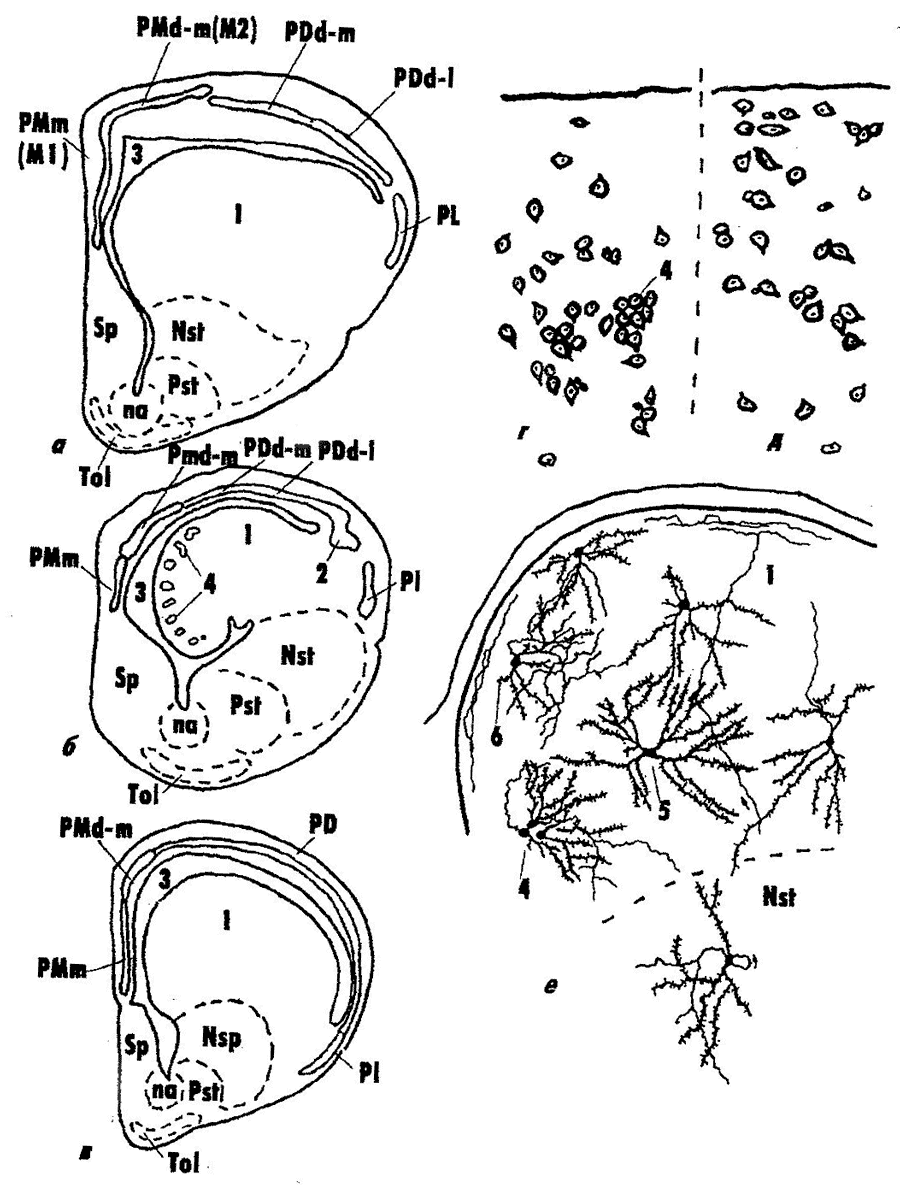 Рис. 5. Структура конечного мозга чешуйчатых рептилий (а), черепах (б) и крокодилов (в), цитоархитектоника дорсо-вентрикулярного края ДВК разного вида и фрагмент нейронной структуры ДВК гадюки (по: Обухов, 1999).
1 – ДВК, 2 – паллиальное утолщение в ДВК, 3 – латеральный мозговой желудочек, 4–6 – разные типы нейронов ДВК, РМm, PMd-m – отделы медиального паллиума, PDd-m, PDd-1 – отделы дорсального паллиума, PL – латеральный паллиум, Nst, Pst – отделы стриатума, Sp – септум, Тol – обонятельный бугорок, na – nucleus accumbens Субпаллиум рептилий включает собственно стриатум и ряд структур вентромедиальной стенки полушария (обонятельный бугорок, септум, амигдалярный комплекс и ряд мелких ядер), которые имеют сходство с соответствующими структурами мозга высших позвоночных. Таким образом, у рептилий происходит качественный скачок в эволюционном развитии ЦНС. Высшие центры деятельности и регуляции работы всей нервной системы и организма переключаются в структуры промежуточного и конечного мозга. На этом этапе эволюции головного мозга позвоночных закладываются все основные предпосылки к дальнейшему развитию высших отделов мозга, включая корковые формации неокортекса млекопитающих и человека. Поэтому изучение структуры и функционирования головного мозга рептилий имеет ключевое значение в понимании процессов эволюции ЦНС позвоночных животных. |
|
Рекомендуемая литература |
|
|