© 2000, Annual Reports of the Zoological Institute RAS.
Olga V. Zherebtsova
Zoological Institute, Russian Academy of Sciences, Universitetskaya nab., 1, St. Petersburg, 199034, Russia
Use of spines for the defence is a characteristic of some representatives of such ancient and conservative groups of mammals as Monotremata (echidnas) and Insectivora (tenrecs, hedgehogs) and also of more advanced Rodentia (porcupines of the New and Old World). A comparative study of defence strategy in different spiny forms of mammals is important for clarifying their adaptive evolution.
The development of spines is connected mainly with the modification of the integument and the subcutaneous muscles. Whole series of investigations on different aspects of structure and functions of these two morpho-functional systems in separate representatives of spiny mammals are known (Michelsson, 1921; Sokolov, 1973; Woods & Howland, 1977; Haffner & Ziswiller, 1983; Vincent & Owers, 1986; Gambaryan & Zherebtsova, 1988; etc.). However, no one has attempted to synthesize such many-sided material in order to reveal and analyse general and specific features of the mammals' defence strategy. This was the major problem in the present research summarizing results of comprehensive comparative analysis of the spiny cover structure and mobility in several groups of mammals.
The fourteen species of mammals (2-5 specimens of each, preserved in 70° alcohol or 5% formalin solution) were studied: echidnas (Tachyglossidae) - Tachyglossus aculeatus; tenrecs (Tenrecidae) - Microgale dobsoni, Tenrec ecaudatus, Hemicentetes semispinosus, Echinops telfairi, Setifer setosus; gymnuras (Erinaceidae, Galericinae) - Hylomys suillus, Echinosorex gymnures; hedgehogs (Erinaceidae, Erinaceinae) - Erinaceus europaeus, E. concolor, Hemiechinus auritus, H. hypomelas, Mesechinus dauuricus; porcupines (Hystricidae) - Hystrix indica. Besides standard histological methods micromorphological techniques were also applied using binocular and electron microscopes and special stain for miozin.
On the basis of the data obtained two major pathways of spiny mammals' specialization were defined. One, which is probably more ancient is related to the development of passive defence strategy. It suggests special spiny armour formation on the dorsal surface of animals and their ability to roll up into a spiny ball in a dangerous situation. Echidnas (Tachyglossus) belonging to Monotremata, hedgehogs (Erinaceus, Hemiechinus, Mesechinus) and hedgehog tenrecs (Setifer, Echinops) belonging to Insectivora use such a way of defence. However, the mechanisms of this strategy differ essentially in these two groups of mammals.
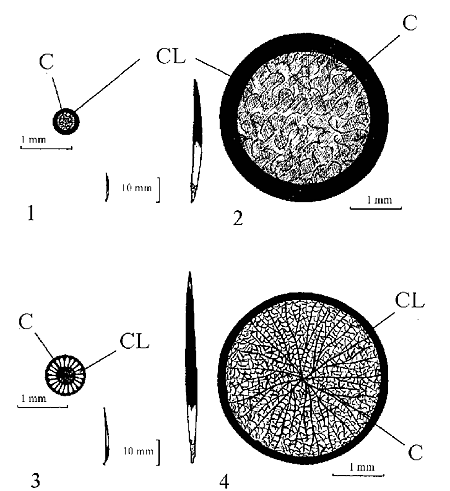
Among forms mentioned above passive defence is less pronounced in hedgehog tenrecs (Eisenberg & Gould, 1970). This is partly attributable to comparatively poor defensive properties of their spines, which are small and slender and are similar to hairs in their internal structure. Continuous cortical layer and extensive region of the core can be seen on the transverse section of tenrec spine (Fig. 1, 1).
The large stout quills of echidna measure up to 60-80 mm in length although they have developed on the basis of a relatively primitive structure, similar to that of tenrecs (Fig. 1, 2). Thus, echidnas have acquired comparatively unwieldy spiny armour.
The spines of hedgehogs are intermediate in dimensions between those of tenrecs and echidnas and differ from them by a more complicated internal structure (Fig. 1, 3). The latter is comprised of special supporting transverse septa and longitudinal stringers in the cortical layer and the core. As a result such features of spines as lightness and rigidity are developed at the same time. Thus, hedgehog armour is less unwieldy in contrast with that of echidna, which is vitally important for hedgehogs characterized by more active mode of life.
The formation of spiny armour in Insectivora has resulted in similar structural modifications of the smooth muscles (mm. arrectores pilorum) in the integument, which are responsible for the spine erection. Mm. arrectores of hedgehog tenrecs and hedgehogs are well developed and consist of a few portions connecting the bulbs of several spines in special complexes, their structure being very similar in both groups (Fig. 2, 1, 2). However, in tenrecs in contrast to hedgehogs the fibres of the subcutaneous muscles (m. cutaneus maximus) are inserted into spine bases. This is evidently related to the communication behaviour of hedgehog tenrecs, which are known to make special sounds by means of spine vibration (Eisenberg & Gould, 1970). It is for these fast movements that the cross-striated subcutaneous muscles are responsible. Unlike insectivorous mammals mm. arrectores are absent completely in the echidna integument, and the function of the large quills mobility is provided by the m. cutaneus maximus only (Fig. 2, 2, 3).
Fig. 1. Scheme of transverse sections of the spines from: 1 - Echinops telfairi; 2 - Tachyglossus aculeatus; 3 - Erinaceus concolor; 4 - Hystrix indica CL - cortical layer; C - core
The development of the continuous spiny cover and the adaptation to passive defence in mammals have resulted also in notable morphological transformations of the subcutaneous muscles. Their functional significance is to serve the entire spiny armour mobility. Hedgehogs are characterized by the most complicated, three-layered structure of the m. cutaneus maximus. In hedgehog tenrecs and echidnas the transformation of the subcutaneous muscles proceeded in the same direction as in hedgehogs, although was not as deep, being confined to two-layered structure.

Fig. 2. Scheme of transverse (a) and tangential (b) sections of a skin in region of the spine ar-mour from: 1 - Erinaceus concolor; 2 - Echinops telfairi; 3 - Tachyglossus aculeatus; 4 - Hystrix indica; 5 - Hemicentetes semispinosus. AP - mm. arrectores pilorum; CM - m. cutaneus maximus; SB - spine bulb; SC - spine case.
Thus, it can be noted the most advanced mechanism of passive defence has undoubtedly developed in hedgehogs. The passive defence in echidnas is also sufficiently effective although it is based upon essentially different and probably more ancient and primitive mechanism.
The other pathway of spiny mammals' specialization is related to the active defence by means of the spines. Such strategy is a characteristic of some tenrecs (Hemicentetes) and porcupines (Hystrix). A common feature of their spines is the unique ability to be easily detached from the animals and to be attached to enemy skin. Different tactics however can be used by these mammals in dangerous situations.
Hemicentetes normally uses the tactics of front defence. These tenrecs possess a special fan of the long slender spines on the crown and try to hit an adversary with their anterior spines (Eisenberg & Gould, 1970). The internal spine structure in Hemicentetes is very similar to that of hedgehog tenrecs (Fig. 1, 1). However, in contrast to the latter roots of spines are placed in special connective tissue sheaths; to their bases simple isolated mm. arrectores pilorum are attached (Fig. 2, 5).
In connection with the main defencive function of the anteriorspines in Hemicentetes its facial m. frontalis is enlarged considerably since it facilitates the mobility of the front part on spiny armour. Meanwhile, the subcutaneous muscle structure is not modified in that case and is very similar to the initial type of the m. cutaneus maximus, which was found in other mammals with skin devoid of spines (Microgale, Hilomys, Echinosorex). The latter have the most simple one-layer structure of the m. cutaneus maximus.
Porcupines (Hystrix) in contrast to Hemicentetes use the tactics of hind or caudal defence. If alarmed and cornered they face away from the adversary, erect their quills, and scatter those by means of rapid tail swing. The large quills of these animals are characterized by more complicated internal structure in contrast to weak and relatively primitive spine structure in Hemicentetes. These quills have the extensive region of the core filled by a special sponge tissue and divided by weakly developed radial septa (Fig. 1, 4). Such quill structure seems to be optimal for striking a single effective blow, but not for standing a long 'siege' as in the case of passive defence.
The mechanism of quill mobility in Hystrix indica is the result of combined action of the well-developed smooth muscles in the integument and the subcutaneous muscles (Fig. 2, 4). Mm. arrectores are joined in linear groups correspondingly to the quills' arrangement along the back. Their contraction results in erection of the separate quills, whereas the simultaneous raising of all linear quill groups at the hind part of the back is the function of the subcutaneous muscles. The latter are well-developed in contrast to those of Hemicentetes and are differenciated into two layers. Similar data are reported for other members of Hystricidae (Woods & Howland, 1977). Thus, porcupines (H. indica) demonstrate more perfect and advanced mechanism of active defence as compared to those of tenrecs (H. semispinosus).
Eisenberg, J.E. & E. Gould. 1970. The tenrecs: a study in mammalian behavior and evolu-tion. Washington, Smithsonian Institution Press. 138 pp.
Gambaryan, P. P. & O.V. Zherebtsova. 1988. Transformation of hypodermic muscles caused by the development of acicular integument in Insectivora (Erinaceidae and Tenre-cidae). Zool. Zh. 67 (1): 79-89 (Communication 1); 67 (2): 189-199 (Communication 2). (In Russian).
Haffner, M. & V. Ziswiler. 1983. Histologische Untersuchungen am lateralen Integument des Igels Erinaceus europaeus. Revue suiss. Zool. 90 (4): 809-816.
Michelsson, G. 1921. Die Hautmusculatur des Igels (Erinaceus europaeus). Gegenbaurs morph. Jb. 51: 147-229.
Sokolov, V.E. 1973. Kozhnyi pokrov mlekopitayushchikh [Skin cover of mammals]. Moskva, Nauka. 487 pp.
Vincent, J.F.V. & P. Owers. 1986. Mechanical design of hedgehog spines and porcupine quills. J. Zool., London (A) 210: 55-75.
Woods, C.A. & E.B. Howland. 1977. The skin musculature of Hystricognath and other se-lected rodents. Zentbl. Vet. Med. comp. Anat. Hist. Embriol. 6: 240-264.