© 2000, Annual Reports of the Zoological Institute RAS.
Vladimir A. Payevsky & Anatoly P. Shapoval
Zoological Institute, Russian Academy of Sciences, Universitetskaya nab., 1, St. Petersburg, 199034, Russia
The number dynamics and natural regulation of avian populations is of considerable importance in the recent ornithological studies. Annual adult survival rates (or mortality rates) along with the productivity rates are the most important demographic parameters. Bird trapping and ringing during breeding as well as during migration is a source of valuable data to study many aspects of birds' life including the demography of populations. The results of ringing are used for the study of survival rates and life expectancy in nature, whereas the numbers of birds caught can be used for the study of population dynamics and age/sex structure of populations. Among population parameters a maximum life span (or simply 'longevity') is the least realistic due to its accidental nature (Gavrilov & Gavrilova, 1992). Nevertheless, observed maximum longevity of vertebrates is commonly used in comparative life-history analyses (Lindstedt & Calder, 1976; Sacher, 1978; Prinzinger, 1979; Golikov, 1985; Payevsky, 1985a; Zammuto, 1986). As demonstrated in North American waterfowl (Krementz et al., 1989), maximum life span was not correlated with the annual survival rate estimates. This seems objectionable in spite of uncertainty of the maximum life span. Theoretically, the survival rate of adult birds and the age structure of a population (and hence the observed value of longevity) should be directly related.
The aims of the present study are as follows: (1) to test the empirical relationships between annual survival rate and maximum life span in birds, and (2) to consider the degree of evenness of age structure in various species of the order Passeriformes.
This report is concerned with bird populations which breed and migrate on the Courish (Curonian) Spit of the Baltic Sea (Kaliningrad Prov., Russia). The birds were trapped and ringed from 1957 by the staff of the Biological Station Rybachy of the Zoological Institute RAS at a permanent field station "Fringilla". The birds were caught in Rybachy-type traps, which resemble Helgoland traps but are larger in size and differ in the method of construction. A total of slightly more than two million birds of 185 species, mainly passerines, were caught and ringed during 40 years, 1957-1996. These birds resulted in ca 7500 recoveries and ca 28000 recaptures. We sexed and aged birds by the methods of Svensson (1970) which were tested and supplemented by us (Vinogradova et al., 1976).
Two sets of data were analyzed: (1) the recaptures of birds from local population which were captured and ringed during re-production period, and (2) the recoveries, i.e. information about rings found at the migratory routes and winter quarters on birds which were ringed during migration. The data for males and fe-males, where possible, were analyzed as different sets. For the following analysis we used the species in which birds can be sexed and aged without the risk of error. The recovery data were analyzed using different models of Brownie et al. (1985) by computer program ESTIMATE. The recovery rates for some species were too sparse for Brownie models to be used, and only the simplest model assuming a constant survival and recovery rate could be applied. The recapture data were analyzed using Jolly-Seber models by program JOLLY (Pollock et al., 1990), and a modified version of the Jolly-Seber model (Buckland & Baillie, 1987).The data for four species fail to fit whatever model, and so the common composite dynamic method was used. Many data on survival were obtained early by us (Payevsky, 1985b; Payevsky et al., 1997).
For the study of age structure of populations we used trapping and ringing data on 21 passerine species. We consider the age structure starting with first year of adult life, i.e. from the yearlings. We used two life span parameters: (1) the age which is observed maximum life span, and (2) the age in which only one per cent or less of birds are still live from the cohort of birds born simultaneously. For both sources of data we used Pearson correlation analysis.
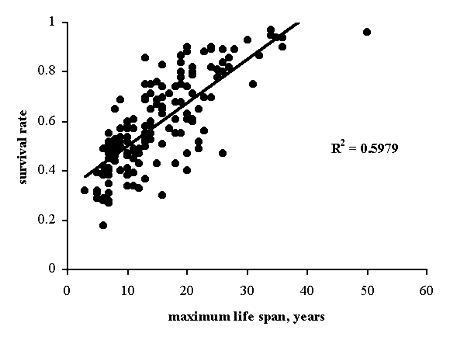
We found that observed longevity of birds is significantly correlated with annual survival rate based on models estimates for either of the life span parameters (r = 0.63 and r = 0.71, df = 29, p < 0.001). We found also significant correlation between the life spans and survival rates obtained from proportion of yearlings (r = 0.77 and r = 0.78, df = 29, p < 0.001). The correlation between maximum longevity and annual survival rates compiled from literature data (156 data sets of 130 species from 13 orders) is also highly significant (r = 0.77, df = 154, p < 0.001, Fig.1).
The range of various demographic parameters for Baltic passerines is presented in Table 1. We found that the coefficient of variation of the species values within the limits of every age class has a significant tendency to increasing from age class of yearlings to age class of ten years old (r = 0.84, df = 8, p = 0.002). The variation is the lowest in age class of two years old and in yearlings. This means that different passerine species have closely similar age structure of the main body of a population, because the birds of one and two years old constitute approximately 80-90 percent of the whole population. This is true equally for males and females.
Fig. 1. Relationship between maximum life spans and survival rates compiled from published data for 130 species of 13 orders.
The findings of our investigation point to the fact that annual survival rate estimates of birds were significantly correlated with observed maximum life spans for either of five data sets. Why do these results differ from earlier ones (Krementz et al., 1989) which indicate that observed maximum life span statistics do not provide a good index to annual survival rate? There is some evidence for the supposition that the reason lies in the specificity of birds group analyzed by Krementz: 16 species of waterfowl with only twofold difference between minimum and maximum life span. We support the opinion of Krementz that observed maximum life span is very sensitive to errors in record keeping, and greatly depends on initial marked sample size, and a length of the study period. Furthermore, as indicated elsewhere (Payevsky, 1974), the life span of birds in captivity is higher than in the nature, and so the observed maximum longevity in the nature is not a "physiologically-determined maximum longevity". We also agree that annual survival rate is a very good life-history parameter for any comparative life-history studies. Just the same, maximum life span values are functions of survival rate, along with other dependencies. That is the reason why even before the analysis there was a priori cause to expect the relationships obtained.
Table 1. Demographic parameters for Baltic passerine species calculated from recoveries and recaptures.
|
Species or species group |
Annual survival rate, % |
Percent of yearlings |
Percent of two-years old birds |
Maximum life span, years |
|
|
from model-based estimates |
from % of yearlings |
||||
|
Motacilla alba |
45.3 |
45.2 |
54.8 |
23.6 |
7 |
|
Anthus pratensis |
38.8 |
38.8 |
61.2 |
19.4 |
5 |
|
Erithacus rubecula |
41.9 |
41.5 |
58.5 |
23.2 |
6 |
|
Turdus spp. |
46.9-53.1 |
45.9-52.6 |
47.4-54.1 |
19.5-24.8 |
6-13 |
|
Hippolais icterina |
40.9 |
40.4 |
59.6 |
24.1 |
10 |
|
Sylvia spp. |
40.1-50.2 |
39.9-46.5 |
53.5-60.1 |
21.2-23.2 |
6-9 |
|
Phylloscopus trochilus |
33.1 |
31.8 |
68.2 |
20.8 |
6 |
|
Regulus regulus |
36.5 |
25.0 |
75.0 |
22.7 |
3 |
|
Parus major |
38.0 |
38.6 |
61.4 |
28.8 |
6 |
|
Emberiza citrinella |
46.8 |
46.5 |
53.5 |
27.4 |
7 |
|
Fringilla spp. |
43.0-56.2 |
52.0-61.8 |
38.2-48.0 |
24.1-29.8 |
9-11 |
|
Carduelis spinus |
48.8 |
53.5 |
46.5 |
30.4 |
12 |
|
Carpodacus erythrinus |
50.4 |
50.2 |
49.8 |
28.1 |
7 |
|
Sturnus vulgaris |
50.1 |
59.3 |
40.7 |
28.1 |
12 |
|
Garrulus glandarius |
49.6 |
49.6 |
50.4 |
20.8 |
8 |
As to age structure patterns in our study, the obtained age distributions for common passerine species provide an insight into the nature of population cycles. There can be little doubt that closer similarity of the age structure reflects similar strategy of reproduction and the recruitment of populations.
We are most grateful to our colleagues at the Biological Station Rybachy for many years of joint efforts in bird trapping and ringing.
Brownie, C., Anderson, D.R., Burnham, K.P. & D.S. Robson. 1985. Statistical inference from band recovery data: a handbook. 2nd ed. Washington, U.S. Fish and Wildlife Service Publ. 305 pp. (Resource Publication 156).
Buckland, S.T. & S.R. Baillie. 1987. Estimating bird survival rates from organised mist-netting programmes. Acta ornithol. 23 (1): 89-100.
Gavrilov, L.A. & N.S. Gavrilova. 1991. Biologiya prodolzhitelnosti zhizni [The biology of life span]. Moskva, Nauka. 280 pp. (In Russian).
Golikov, A.N. 1985. On a body size change and longevity of organisms during the evolution. Zh. obshch. Biol. 46 (6): 771-777 (In Russian).
Krementz, D.G., Sauer, J.R. & J.D. Nichols. 1989. Model-based estimates of annual survival rate are preferable to observed maximum lifespan statistics for use in comparative life-history studies. Oikos 56 (2): 203-208.
Lindstedt, S.L. & W.A. Calder. 1976. Body size and longevity in birds. Condor 78 (1): 91-145.
Payevsky, V.A. 1974. Longevity and annual mortality in birds. Trudy Zool. Inst., Leningrad 55: 142-185. (In Russian).
Payevsky, V.A. 1985a. Body mass and longevity in birds. Vest. Zool. (2): 70-72. (In Russian).
Payevsky, V.A. 1985b. Demografiya ptits [Demography of birds]. Leningrad, Nauka. 285 pp. (In Russian).
Payevsky, V.A., Vysotsky, V.G., Yefremov, V.D., Markovets, M.Y. & A.P. Shapoval. 1997. Sex-specific survival rates in birds. Zh. obshch. Biol. 58 (6): 5-20.
Pollock, K.H., Nichols, J.D., Brownie, C. & J.E. Hines. 1990. Statistical inference for capture-recapture experiments. Washington, The Wildlife Society Publ. 97 pp. (Wildlife Monograph 107).
Prinzinger, R. 1979. Lebensalter und Gesamtenenergieproduction beim Vögel. J. Ornithol. Lpz. 120 (1): 103-105.
Sacher, G.A. 1978. Longevity and aging in vertebrate evolution. BioScience 28 (8): 497-501.
Svensson, L. 1970. Identification Guide to European Passerines. Stockholm, Naturhist. Riksmuseet. 152 pp.
Vinogradova, N.V., Dolnik, V.R., Yefremov, V.D. & V.A. Payevsky. 1976. Opredeleniye pola i vozrasta vorobyinych ptits fauny SSSR [Sex and age determination of the Passerine birds of the USSR]. Moskva, Nauka. 192 pp. (In Russian).
Zammuto, R.M. 1986. Life histories of birds: clutch size, longevity, and body mass among North American game birds. Can. J. Zool. 64 (12): 2739-2749.