© 2000, Annual Reports of the Zoological Institute RAS.
To the knowledge of larvae of leafhoppers of the subfamily Deltocephalinae
(Homoptera, Cicadellidae)
Dmitry A. Dmitriev
Zoological Institute, Russian Academy of Sciences, Universitetskaya nab., 1, St. Petersburg, 199034, Russia
Although Cicadina is a well studied group, their larvae remain insufficiently known. This concerns particularly the larvae of leafhoppers, because they are rather uniform and their identification is difficult. At present there are only a few works with the descriptions of larvae of
Cicadellidae, keys for their identification and comparative morphological data (Walter, 1975, 1978; Wilson, 1978, 1981, 1983; Vilbaste, 1982; Dmitriev, 1999).
The significance of study of larvae is evident. Above all this could permit us to differentiate species with only their larvae in collections. The larvae give also complementary characters for the description of higher taxonomic groups. Eventually all this could help to comprehend the phylogeny of the group.
The system of Cicadina is based mainly on characters of male genitalia. However this method cannot be used for larvae. Previous authors used scanty set of characters for the description of larvae, mainly such as the pattern and chaetotaxy of the abdomen, and also some other striking characters. In some cases, however they are not enough for the description of suprageneric and even generic
taxa.
The present investigation is confined to a study of leafhoppers' nymphs of the subfamily Deltocephalinae. It is the largest and the most diverse subfamily in the family Cicadellidae. Detailed descriptions of larvae of more than 150 species have been composed. These species represent all tribes of the subfamily from the Palaearctic region.
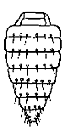
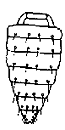
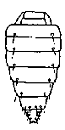
Of the characters used in descriptions chaetotaxy of the abdomen should be noted especially. It is characterised by regressive evolution. Ten principal patterns of chaetotaxy are presented in the Table.
The 1st type of chaetotaxy is numerous setae covering the whole body. There are also some macrosetae among them. They arise from hind margins and from hind angles of abdominal tergites. This type of chaetotaxy is observed in representatives of 3 tribes: Drabescini, Tetartostylini and Macrostelini. In all other cases setae arise from hind margins of tergites III-VIII, from hind angles of tergites VII-VIII, and from pygofer, sometimes the chaetotaxy is more or less reduced.
Table. Abdominal chaetotaxy of leafhoppers of the subfamily Deltocephalinae
|
Tribes |

1 |

2 |
3 |
4 |

5 |
6 |

7 |

8 |

9 |

10 |
|
Drabescini |
+ |
|
|
|
|
|
|
|
|
|
|
Macrostelini |
+ |
|
|
|
|
+ |
+ |
+ |
+ |
|
|
Hecalini |
|
+ |
|
|
|
|
+ |
|
|
|
|
Opsiini |
|
+ |
+ |
+ |
+ |
|
|
+ |
|
|
|
Athysanini |
|
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
|
Scaphytopiini |
|
|
+ |
+ |
+ |
|
|
|
|
|
|
Doraturini |
|
|
+ |
+ |
|
|
|
|
+ |
+ |
|
Paralimnini |
|
|
+ |
+ |
|
|
|
|
|
|
|
Goniagnathini |
|
|
|
+ |
+ |
|
|
|
|
|
|
Stirellini |
|
|
|
+ |
+ |
|
+ |
|
|
|
|
Deltocephalini |
|
|
|
? |
+ |
|
|
|
|
|
|
Selenocephalini |
|
|
|
|
+ |
|
|
|
|
|
|
Tetartostylini |
|
|
|
|
+ |
|
|
|
|
|
|
Fieberilini |
|
|
|
|
|
+ |
|
|
|
|
|
Grypotini |
|
|
|
|
|
|
+ |
|
|
|
|
Limotetigini |
|
|
|
|
|
|
|
+ |
+ |
|
Notes: explanations are in the text.
The 2nd type - numerous setae on each tergite. 8-14 setae arise from fore tergites, the number of setae usually decreases backwards. It is found in Opsiini and
Athysanini.
The 3rd type - 6 setae on each tergite. However 6 setae on tergites III-VI are not homologous to 6 setae on tergites VII-VIII, where
intermediate setae between medial and lateral setae are absent but there are
additional ones in the hind angles.
The 4th type of chaetotaxy is the most common one in the subfamily: 4 longitudinal rows of setae and additional ones in the hind angles.
The 5th type may be defined as a decrease in the number of setae on tergite VII and often VIII.
The 6th type. Median rows of setae are absent, there are only lateral ones. This is found in Fieberilini and in species of
Platymetopius (Athysanini).
The 7-9th types are the stages of further evolution of chaetotaxy. Setae arise from tergites VII-VIII, from tergite VIII, and finally from pygofer only.
The 10th type is the total absence of abdominal setae. It is found only in
Chiasmus (Doraturini).
It should be noted that 6 longitudinal rows of setae or 4 rows with additional setae in the hind angles of tergites VII-VIII take place in many tribes. This provides evidence of parallel changes of this character in the different tribes.
Genera Eupelix, Dorycephalus and Paradorydium are placed by some authors in the subfamily Deltocephalinae as three separate tribes
(Anufriev & Emeljanov, 1988; Emeljanov, 1999). However they are undoubtedly more closely related to each other than to the other representatives of the subfamily. This implies that they should be considered as a
separate subfamily Eupelicinae. Furthermore it is worth noting that Paradorydium is more closely related to
Eupelix than to Dorycephalus. Their synapomorphy is the presence of platellae on the apex of the hind tibia, which is retained also in imago (Eupelix) or turns into common setae (Paradorydium). At the same time
Dorycephalus has its own apomorphies: longitudinal grooves on postclypeus and platellae in the
anteroventral row on the first segment of the hind tarsus.
At present it is possible to suggest a preliminary key to Palaearctic tribes of leafhoppers of the subfamily Deltocephalinae based on
characters of nymphs:
A preliminary key to tribes of the subfamily Deltocephalinae
1(6). Whole body covered with numerous setae.
2(3). Pygofer deeply incised (more than 2/3 of its length), the bases of the lobes widely apart from one another . . . . . . . .
Drabescini
3(2). Pygofer incised less than 1/2 its length, the bases of lobes closed.
4(5). Vertex medially longer than laterally. Pattern consist of black dots . . . . . . . .
Tetartostylini
5(4). Vertex medially as long as laterally. Pattern consist of characteristic black and yellow patches . . . . . . . .
Macrostelini (part)
6(1). Setae arise only from hind margins of abdominal tergites or absent.
7(10). Pygofer strongly elongated (more than 2.5 times as long as tergite VIII).
8(9). Abdomen with 4-6 longitudinal rows of setae . . . . . . . .
Scaphytopiini
9(8). Abdomen with 2 lateral rows of setae . . . . . . . . Fieberilini
10(7). Pygofer less than 2 times as long as tergite VIII.
11(12). Abdomen with pair of additional lateral carinae (besides carinae separating dorsal parts of tergites and ventral laterotergites) . . . . . . . .
Selenocephalini
12(11). Abdomen without additional carinae.
13(14). Head spade-shaped, its sides usually foliaceously flattened . . . . . . . .
Hecalini
14(13). Head not spade-shaped.
15(18). Frons proceeds to face surface, postclypeus does not reach vertex.
16(17). Abdomen with 2 setae on tergite VII and 6 on VIII. Anal tube with 6 pairs of setae . . . . . . . .
Grypotini
17(16). Other type of abdominal chaetotaxy. Anal tube with 4 pairs of setae . . . . . . . .
Macrostelini (part)
18(15). Frons on vertex surface; postclypeus proceeds to vertex surface as two patches on each side of frons.
19(22). Dorsal rows of setae on metatibiae consist of 6-7 macrosetae.
20(21). First seta on apex of first segment of hind tarsus is common.
Abdomen with 4 longitudinal rows of setae . . . . . . . . Deltocephalini
21(20). First seta on apex of first segment of hind tarsus is platella.
Abdomen with setae on tergite VIII and pygofer or on pygofer only . . . . . . . .
Limotettigini (Including genera: Limotettix, Ophiola, Anoterrostemma.)
22(19). Dorsal rows of setae on metatibiae consist of 8-11 macrosetae.
23(28). Anteclypeus strongly widening towards apex (more than 1.2 times).
24(25). Vertex with cavity before apex. Metafemur with 2+2+1+1 setae at apex . . . . . . . .
Opsiini (part)
25(24). Vertex flat or convex. Metafemur with 2+2+1 setae at apex.
26(27). Postclypeus nearly as long as wide. Genae with straight margins in lower part and deep cavities at eyes. First segment of hind tarsus with 4 platellae and common seta at apex . . . . . . . .
Goniagnathini
27(26). Postclypeus longer than wide. Lateral margins of genae evenly rounded, with weak cavities at eyes. First segment of hind tarsus only with platellae at apex . . . . . . . .
Athysanini (part)
28(23). Anteclypeus parallel-sided or slightly widening or narrowing towards apex.
29(30). Abdomen with 4 longitudinal rows of setae, tergite VII with 2 setae, VIII with 4-6 setae. Setae may be very short, almost
indistinct, in this case first segment of hind tarsus with platellae in anteroventral row. Fore margin of vertex rounded . . . . . . . .
Opsiini (part)
30(29). Other type of chaetotaxy. When chaetotaxy is such as above, fore margin of vertex is angled.
31(32). Pygofer flattened laterally, almost without dorsal cavity. Wing pads strongly reduced, apices of fore wing pads do not reach apices of hind wing pads . . . . . . . .
Opsiini (part)
32(31). Pygofer not flattened, deeply incised dorsally. Wing pads more or less developed.
33(36). Metafemur with 2+1 setae at apex. Face longer than wide.
34(35). Abdomen bald . . . . . . . . Doraturini (part)
35(34). Abdomen with 4 complete or incomplete rows of setae . . . . . . . .
Stirellini
36(33). Metafemur with 2+2+1 setae at apex.
37(38). Abdomen with setae on tergites VII-VIII only, or with more than 4 longitudinal rows of setae, if there are 4 rows, than tergites VII-VIII not with 6 setae . . . . . . . .
Athysanini (part)
38(37). Tergites III-VI with 4 or 6 setae, VII-VIII with 6 setae, or abdomen bald, with setae on pygofer only.
39(40). Lorum not bordered or less than 1/4 of its length bordered on postclypeus; anteclypeus parallel-sided or narrowing towards apex. Fore margin of vertex usually angled. Sides of pronotum without carinae. First segment of hind tarsus with platellae and common seta at apex . . . . . . . .
Paralimnini
40(39). 1/3-1/2 of inner margin of lorum bordered on postclypeus; ante-clypeus parallel-sided or somewhat widening towards apex. Fore margin of vertex usually rounded. Sides of pronotum usually with weak carinae. First segment of hind tarsus only with platellae at apex.
41(42). Anteclypeus parallel-sided; inner angle of lorum falls on junction of anteclypeus and postclypeus and divides inner edge of lorum into two unequal parts. Face usually flattened, postclypeus and vertex often with cavities. Wing pads usually short. Male gonapophyses usually as long as wide, with widely rounded apices . . . . . . . .
Doraturini
42(41). Anteclypeus usually somewhat widened towards apex, narrowed at the base, and its lateral edge S-shaped; inner angle of lorum falls on the most narrow part of anteclypeus and divides inner edge into two equal parts. Face and vertex without cavities. Male
gonapophyses usually longer than wide at the base, with pointed and somewhat elongated apices, so their lateral edges S-shaped . . . . . . . .
Athysanini (part)
Acknowledgements
I am thankful to Prof. A.F. Emeljanov, my scientific adviser and N.A. Vassilieva for
linguistic advice.
References
Anufriev, G.A. & A.F. Emeljanov. 1988. Suborder Cicadinea (Auchenorrhyncha). In:
Opredelitel' nasekomykh Dal'nego Vostoka SSSR [Keys to the Insects of the Far East of the USSR]. Vol. 2. pp. 12-495. Leningrad, Nauka. (In Russian).
Dmitriev, D.A. 1999. Larvae of the European species of Elymana DeLong
(Homoptera: Cicadellidae). Zoosyst. Ross. 8(1): 77-78.
Emeljanov, A.F. 1999. A key to genera of the subfamily Deltocephalinae s.l. (Homoptera, Cicadellidae) from Kazakhstan, Middle Asia, and Mongolia with description of new
genera and subgenera. Ent. Rev. 79 (5): 547-562.
Vilbaste, J. 1982. Preliminary key for the identification on the nymphs of North European Homoptera Cicadina. II. Cicadelloidea. Ann. zool. fenn. 19 (1): 1-20.
Walter, S. 1975. Larvenformen mitteleuropaischer Euscelinen (Homoptera, Auchenorhyncha). Zool. Jb. (Syst. Abt. 3) 102 (2): 241-302.
Walter, S. 1978. Larvenformen mitteleuropaischer Euscelinen (Homoptera, Auchenorhyncha). Teil II. Zool. Jb. (Syst. Abt. 3) 105 (1): 102-130.
Wilson, M.R. 1978. Descriptions and key to the genera of the nymphs of British woodland Typhlocybinae (Homoptera). Syst. Ent. 3 (1): 75-90.
Wilson, M.R. 1981. Identification of European Iassus species (Homoptera: Cicadellidae) with one species new to Britain. Syst. Ent. 6: 115-118.
Wilson, M.R. 1983. The nymphal stages of some Auchenorrhyncha associated with rice in south East Asia. In: Proceedings of the 1st International Workshop on Leafhoppers and Planthoppers of Economic Importance. pp. 121-134. London, Commonwealth Institute of Entomology
Publ.