
IISN 1608-4195
А.В. БАЛУШКИН, О.Е. БОГОДИСТ, К.Э. СКУРА, О.Ю. ТЕРЕЩУК
САНКТ-ПЕТЕРБУРГ, 2002
Главный редактор – директор Зоологического института РАН А.Ф. Алимов
Редакционная коллегия:
В.В. Хлебович, Я.И. Старобогатов, С.Д. Гребельный,
Т.А. Асанович, Ю.С. Балашов, В.Я. Бергер, И.С. Даревский,
В.Р. Дольник, С.Ю. Кузнецов, А.В. Горохов
Редакторы выпуска:
Б.А. Шейко, В.В. Федоров
Рецензенты:
Г.П. Руденко, Н.В. Максимович
В номере приведены морфологические характеристики видов и отдельных популяций бычковидных нототений рода Gobionotothen Balushkin, мраморной нототении Notothenia rossii Richardson островов Кергелен (семейство Nototheniidae) и парусовидной бородатки Histiodraco velifer (Regan) (семейство Artedidraconidae), основанные на материалах российских, польских и немецких экспедиций в Антарктику. Дана оценка общего сходства популяций зеленой нототении G. gibberifrons (Lonnberg) из различных районов моря Скотия и парусовидной бородатки из морей Дейвиса, Космонавтов, Росса, Содружества и Уэдделла с помощью фенетических методов. Обсуждается феномен итеративной эволюции, имевший место при диверсификации видов Gobionotothen.
Настоящее издание осуществлено при поддержке программы ФЦП «Изучение и исследование Антарктики», проект No16 «Проведение комплексного изучения антарктической биоты», ФЦП «Интеграция» (проект А0144) и грантов Российского фонда фундаментальных исследований No 00-04-48838 «Рыбы Южного океана (систематика, эволюция, зоогео-графия)», No 00-15-07794 «Петербургская ихтиологическая школа».
Издание предназначено для публикации отдельных работ Зоологического Института на русском и английском языках. Публикуется без определенной периодичности и распространяется по заявкам.
© Зоологический институт РАН, 2002
© А.В. Балушкин, О.Е. Богодист, К.Э. Скура, О.Ю. Терещук, 2002
ЗИН РАН, 199034, СПб., Университетская наб., 1
Тел.: (812) 3280311 Факс: (812) 3282941 E-mail: admin@zin.ru
К настоящему времени фауна рыб Южного океана насчитывает 367 видов, относящихся к 59 семействам (Balushkin, Fedorov, 2001; Балушкин, Федоров, 2002). Доминирующее место в ней по численности и таксономическому разнообразию занимает подотряд нототениоидных рыб Notothenioidei, объединяющий 8 семейств, 48 родов и 139 видов (Balushkin, 2000). Грандиозные геологические перестройки земной коры, структуры океанических вод и климата в южном полушарии в кайнозое создали условия для уникального в истории рыб феномена возникновения таксона высокого ранга, сформировавшего в ходе длительной изоляции практически монофилетический ихтиоцен на огромной акватории Мирового океана. Этим обстоятельством во многом объясняется повышенный интерес ученых различных стран мира к данной группе животных для решения многих общих проблем биологии и исторической биогеографии.
Холодный климат, суровый ледовый режим и удаленность от мест с развитой цивилизацией способствовали сохранению в неприкосновенности биотопов антарктических вод вплоть до середины 70 гг. прошлого века, т.е. до времени начала крупномасштабного тралового промысла в Южном океане. Интенсивность промысла была столь велика, что к концу столетия было выловлено около 3 млн. т рыб, в первую очередь таких массовых видов, как мраморная нототения Notothenia rossii Richardson, серая нототения, или сквама, Lepidonotothen squamifrons (Gunther), зеленая нототения Gobionotothen gibberifrons (Lonnberg), патагонский клыкач Dissostichus eleginoides (Smitt) и ледяная рыба Champsocephalus gunnari (Lonnberg) (Шуст, 1998). К сожалению, научные наблюдения в эти годы явно отставали от темпов промышленного освоения, и, как следствие этого, сегодняшние наши знания даже о промысловых рыбах Антарктики бедны и во многом отрывочны. После введения серьезных запретительных мер на лов рыбы Комиссией по сохранению морских живых ресурсов Антарктики (АНТКОМ) число промысловых экспедиций в Антарктику резко сократилось. Из-за неблагополучной экономической ситуации в нашей стране были практически свернуты также научные программы изучения антарктической морской биоты. Поэтому большое значение сейчас приобретает анализ и обобщение ретроспективных данных, касающихся, в частности, вопросов видового полиморфизма и популяционной структуры антарктических рыб.
В этом номере приведены результаты сравнительно-морфологического изучения трех видов нототениоидных рыб, проводившегося А.В. Балушкиным в разные годы совместно с польским ихтиологом К.Э. Скурой на Морской станции Гданьского университета (г. Хель, Польша) и со студентами кафедры ихтиологии и гидробиологии Санкт-Петербургского университета О.Ю. Терещук и О.Е. Богодист.
МАТЕРИАЛ И МЕТОДЫ
При изучении бычковидных нототений были использованы материалы, собранные в экспедициях на НИС "Профессор Сидлецки" (1978-1979 гг.), "Гемини" (1976-1977 гг.) (данные К. Э. Скуры), "Гижига" (1984-1985 гг.) (данные А.В. Балушкина), а также при обработке коллекционных материалов других антарктических экспедиций.
Gobionotothen gibberifrons. Район антарктического полуострова: «Проф. Сидлецки", тр. 92, 93, 107 и 108; "Гижига", тр. 136. Число экз.: 306 (vert.), 100 (Р), 80 (D1, D2, A), 40 (ULL, MLL), 20 (для остальных признаков). Южные Шетландские о-ва: «Проф. Сидлецки", тр. 58, 94, 107 и 108; "Гижига", тр. 107. Число экз.: 500 (vert.), 160 (P), 140 (D1, D2, A), 40 (ULL, MLL), 20 (для остальных признаков). Южные Оркнейские о-ва: "Гемини", тр. 61, 62, 63 и 64; «Проф. Сидлецки", тр. 23 и 127; "Гижига", тр. 256, 265 и 281. Число экз.: 297 (vert.), 140 (P), 110 (D1, D2, A), 60 (ULL, MLL), 30 (для остальных признаков). Остров Ю. Георгия: "Гемини", тр. 13, 20, 23; «Проф. Сидлецки", тр. 115, 136, 138, 139; "Зунд", тр. 92, 98. Число экз.: 295 (vert.), 205 (P), 195 (D1, D2, A), 20 (ULL, MLL), 10 (для остальных признаков).
G. barsukovi. "Обь", 1957 г., ст. 276; "Аэлита", 1969 г., тр. 59, 77, 134; "Аэлита", 1970 г., ст. 280; "Скиф", 1969 г., тр. 297; "Скиф", 1970 г., тр. 97, 109; "Кара-Даг", 1971 г., тр. 200; "Кара-Даг", 1972 г., ст. 543; "Фиолент", 1972 г., тр. без номера. Число экз.: 38 (P, ULL, MLL), 19 (для остальных признаков).
G. angustifrons. "Советская Украина", 1965 г., ст. 717; "Академик Книпович", 1981 г., тр. 102. Число экз.: 62 (P, ULL, MLL), 37 (vert.), 31 (для остальных признаков).
G. marionensis. "Аэлита", 1968 г., 2 трала (без номеров). Число экз.: 8 (P, ULL, MLL), 4 (для остальных признаков).
G. acuta. "Challenger", Kerguelen. Голотип BNHM 1879.5.14.360.
Более подробную информацию о перечисленных выше тралениях можно найти в следующих работах: Балушкин, 1986, 1991; Воскобойникова, Балушкин, 1987; Sosinski, Skora, 1988.
Материалом для изучения мраморной нототении послужили сборы российских антарктических экспедиций 1970–1971 гг. на НПС “Скиф” и “Кара-Даг”, хранящиеся в коллекциях Зоологического института РАН.
Notothenia rossii ЗИН No40244, “Скиф”, о. Кергелен, 48°08'1 ю.ш., 70°18.5' в.д., тр. 529, глуб. 140–144 м, 16.03.1970 г., колл. Н.В. Кононов (4 экз.); No 42495, “Кара-Даг”, о. Кергелен, ст. 200, глуб. 290–300 м, 11.09.1971 г., колл. Ю.Н. Щербачев и Э.Э. Мейснер (1 экз.); No 44311, “Скиф”, о. Кергелен, тр. 512, 13.03.1970 г., колл. Н.В. Кононов (4 экз.); No 45013, “Скиф”, о. Кергелен, 48°01'8 ю.ш., 70°27'6 в.д., ст. 1047/136, тр. 97, глуб. 146 м, 25.12.1970 г., колл. А.Ф. Пушкин (2 экз.); No48554, “Скиф”, о. Кергелен, 49°54'5 ю.ш., 70°39'2 в.д., ст. 997/86, тр. 63, глуб. 263–275 м, 14.12.1970 г., колл. А.Ф. Пушкин (1экз.); No48555, “Скиф”, о. Кергелен, 47°20'7 ю.ш., 70°32'2 в.д., ст. 1019/108, тр.68, глуб. 162 м, 20.12.1970 г., колл. А.Ф. Пушкин (1 экз.); No 48556, “Скиф”, о. Кергелен, 48°02'6 ю.ш., 70°37'1 в.д., ст. 1057/146, тр 106, глуб. 147–149 м, 26.12.1970 г., колл. А.Ф. Пушкин (1 экз.); No48557, “Скиф”, о. Кергелен, 47°59'9 ю. ш., 70°16'9 в.д., ст. 1123/212, тр. 140, глуб. 150 м, 11.01.1971 г., колл. А.Ф. Пушкин (3 экз.); No48558, “Скиф”, о. Кергелен, 48°24' ю.ш., 70°05'4 в.д., ст. 1144/233, тр. 152, 13.01.1971 г., колл. А.Ф. Пушкин (1 экз.); No 48559, “Скиф”, о. Кергелен, 48°27'6 ю.ш., 70°02'16 в.д., ст. 1156/245, тр. 164; глуб. 133–134 м, 15.01.1971 г., колл. А.Ф. Пушкин (1 экз.); No52833, “Кара-Даг”, ст. 283, тр. 1625, глуб. 250–280 м, 29.09.1971 г., колл. Ю.Н. Щербачев и Э.Э. Мейснер (1 экз.).
Материалом для изучения парусовидной бородатки послужили коллекции Зоологического института РАН (ЗИН) и Зоологического музея Гамбургского университета (ISH).
Histiodraco velifer. ЗИН No37500, "Обь", море Содружества, 66°48,2' ю.ш., 63°50–25' в.д., ст. 198а, 31.01.1957 г. (1 экз. 123 мм SL); ЗИН No46459, "Мыс Юноны", море Росса, о. Росса, тр. 20, глуб. 260 м, 9.02.1982 г. (1 экз. 159 мм SL); ЗИН No 52132, "Чатыр-Даг", море Содружества, тр. 140, 7.03.1979 г. (2 экз. 117 и 157 мм SL); ЗИН No52133, "Чатыр-Даг", море Моусона, тр. 52 (53) (1 экз. 158 мм SL); ЗИН No52135, "Мыс Юноны", море Росса, о. Франклина, тр. 70, глуб. 30–80 м, 20.02.1993 г. (3 экз. 119–157 мм SL); ЗИН No52135, "Звезда Азова", море Космонавтов, 67°31' ю.ш., 43°57' в.д., тр. 79, 3.03.1988 г. (9 экз. 117–152 мм SL); ISH 84–1985, "Polarstern", рейс 06, море Уэдделла, 77°28.9' ю.ш., 41°26.6' з.д., ст. 299, трал 07, глуб. 667 м, 5.02.1985 г. (1 экз. 122 мм SL); ISH 196–1981, "Polarsirkel", море Уэдделла, 70°31.9' ю.ш., 07°22.9' з.д., ст. 143/81, глуб. 280 м, 27.01.1981 г. (1 экз. 142 мм SL); ISH 69–1991, "Polarstern", море Уэдделла, 70°24.1' ю.ш., 06°07.6' з.д., ст. 220, глуб. 118–126 м, 13.03.1991 г. (1 экз. 145 мм SL); ISH 53–1984, "Polarstern", море Уэдделла, 76°56.7' ю.ш., 49°44.0' з.д., ст. 474, глуб. 224 м, 14.02.1984 г. (1 экз. 127 мм SL); ISH 46–1984, "Polarstern", море Уэдделла, 75°46.1' ю.ш., 56°50.8' з.д., ст. 417, глуб. 348 м, 5.02.1984 г. (1 экз. 97 мм SL); ISH 52–1983, "Polarstern", море Уэдделла, 70°30.3' ю.ш., 08°04.0' з.д., ст. 220, глуб. 261–263 м, 1.02.1983 г. (1 экз. 115 мм SL), ISH 46–1983, "Polarstern", море Уэдделла, 75°38.6' ю.ш., 27°20.5' з.д., ст. 196, глуб. 288–287 м, 23.02.1983 г. (1 экз. 127 мм SL).
Методика измерений. SL – стандартная длина, от вершины верхней челюсти до конца гипуралий; H – высота тела, от первого луча анального плавника до основания лучей 2-го спинного плавника, aD – антедорсальное расстояние (aD1 – от вершины верхней челюсти до 1-го луча первого спинного плавника; aD2 – от вершины верхней челюсти до 1-го луча второго спинного плавника); HD1(hD1), HD2(hD2) – наибольшая высота соответственно первого и второго спинных плавников; aA – антеанальное расстояние (aA1 – от вершины верхней челюсти до первого луча анального плавника, aA2 – от вершины нижней челюсти до 1-го луча анального плавника; aВr – антебранхиальное расстояние, наименьшее расстояние от вершины нижней челюсти до заднего края межжаберной перепонки; LP(lP) – длина грудного плавника, от основания 1-го луча до конца самых длинных лучей; LV(lV) – длина брюшного плавника, от основания первого луча до конца самого длинного луча; LVA(lVA) – вентроанальное расстояние, от основания первого луча брюшного плавника до первого луча анального плавника; Lср(lcp) – длина хвостового стебля, от точки пересечения перпендикуляра, восстановленного от последнего луча анального плавника, с горизонтальной осью тела до конца гипуралий; Hср(hcp) – высота хвостового стебля, на уровне середины расстояния между последним лучом второго спинного плавника и первым краевым лучом хвостового плавника; LC(lc) – длина головы (lc1 – от вершины верхней челюсти до заднего костного края жаберной крышки; lc2 – от вершины верхней челюсти до средней поры супратемпоральной комиссуры); Wc – ширина головы, расстояние между предкрышками обеих сторон головы; Hc(hc) – высота головы, на уровне основания первого луча брюшного плавника; Ho(ho) – высота головы через середину глаза; aO – длина рыла, от вершины верхней челюсти до переднего края глазницы по прямой, проходящей через ноздрю; О(o) – продольный диаметр орбиты; Lmx(lmx) – длина верхней челюсти; Lmd(lmd) – длина нижней челюсти; iO(io) – межглазничное расстояние (пространство), между наружными краями frontalia; ID(D1), IID(D2), P, А – число лучей в первом и втором спинных, грудном и анальном плавниках; ULL(ULl), MLL(MLl) – число трубчатых чешуй в дорсальной и медиальной (срединной) боковых линиях; sp.br. – число жаберных тычинок (sp.br.ant. – во внешнем ряду; sp.br.post. – во внутреннем ряду первой жаберной дуги; Sq – число поперечных рядов чешуй туловища; vert. – общее число позвонков. Эта методика основана на ранее опубликованной (Балушкин, 1976), в которую внесен ряд дополнений и уточнений.
Измерения тела отнесены к стандартной длине (SL), измерение головы – к ее длине (lc1). Для выборок вычисляли размах варьирования признака lim, среднее арифметическое значение признака x, ошибка выборочной средней mx, среднее квадратичное отклонение s. Для сравнения выборок мраморной нототении был использован коэффициент различия CD Майра.
Расчеты фенетических дистанций основаны на стандартизированных (нормированных) средних значениях признаков. В качестве мер различия использовано евклидово расстояние с применением различных алгоритмов (методов) кластеризации: Single Linkage (nearest neighbor), Complete Linkage (farthest neighbor), Centroid Linkage, Average Linkage, Median Linkage и Ward MinimumVariance Methods.
Авторы признательны Б.А. Шейко и В.В. Федорову за редактирование рукописи, коллекторам К.-Г. Коку, Н.В. Кононову, Э.Э. Мейснеру, А.Ф. Пушкину, П. Храповой, М. Штеманну, Ю.Н. Щербачеву и другим участ-никам экспедиций за сбор материала для исследования, А.О. Юрцевой и Т.Б. Николаевой за изготовление оригинал-макета этого номера известий ЗИН РАН. А.В. Балушкин благодарен проф. Хорсту Вилкенсу за любезное приглашение посетить Зоологический институт Гамбургского университета для изучения коллекций антарктических рыб и ассистенту Г. Шульце за помощь в работе и изготовление рентгеноснимков.
ОБЩЕЕ СХОДСТВО И ОСОБЕННОСТИ ЭВОЛЮЦИИ БЫЧКОВИДНЫХ НОТОТЕНИЙ РОДА GOBIONOTOTHEN BALUSHKIN (NOTOTHENIIDAE) ЮЖНОГО ОКЕАНА
K роду зеленых или бычковидных нототений Gobionotothen относятся 5 видов – G. marionensis (Gunther, 1880), G. angustifrons (Fischer, 1885), G. acuta (Gunther, 1880), G. barsukovi (Balushkin, 1991) и G. gibberifrons (Lonnberg, 1905), населяющих западноантарктические острова моря Скотия от скал Шаг на севере до Антаpктического полуострова на юге и острова индоокеанского сектора Антарктики (Пpинс-Эдуаpд, Кpозе, Кеpгелен и Х¨pд) (Балушкин, 1991; Balushkin, 2000).
Интенсивные научные и научно-промысловые исследования второй половины ХХ века поставили ряд проблем, касающихся морфологической дифференциации видов и отдельных популяций антарктических рыб. Одна из таких проблем, вызвавших дискуссию в литературе, связана с анализом морфологической обособленности популяций бычковидных нототений в различных регионах Южного океана и оценкой их таксономического статуса (Andersen, Hureau, 1979; Andersen, 1984; Балушкин, 1986; Воскобойникова, Балушкин, 1987; Gon, Klages, 1988; Balushkin, 1989; DeWitt et al., 1990; Балушкин, 1991, 1992). Цель настоящей работы – обобщить имеющиеся у каждого из авторов морфометрические данные по видам и отдельным популяциям зеленых нототений, собранных в российских и польских экспедициях в Антарктике, провести анализ их сходства с помощью фенетических методов и обсудить характер и возможные пути эволюции рода. Использование фенетических методов было продиктовано тем, что проведенный ранее биометрический анализ отдельных признаков не выявил статистически значимых отличий между выборками зеленой нототении из разных районов видового ареала (Балушкин, 1986).
Расчеты фенетических дистанций выполнены по средним значениям 15 пластических (aD1, aD2, aA1, LP, LV, Lcp, Hcp, Lc1, Wc, Ho, aO, O, Lmx, Lmd, iO) и 9 меристических признаков (ID, IID, A, P, ULL, MLL, sp.br.ant., sp.br. inn., vert.). У G. gibberifrons средние значения были подсчитаны раздельно для каждой выборки из 4 районов ареала (о-ва Южная Георгия, Южные Оркнейские, Южные Шетландские и северная оконечность Антарктического полуострова), что позволило сравнить уровень видовых и популяционных различий.
Общее представление о величине фенетической обособленности отдельных выборок зеленой нототении G. gibberifrons из различных районов Западной Антарктики и всех других видов рода бычковидных нототений по комплексу из 24 счетных и пластических признаков дают дендрограммы, представленные на рис. 1. Как это можно было предполагать a priori, среди всех включенных в анализ О.Т.Е. наиболее близкими друг другу оказались выборки G. gibberifrons: они оказались сгруппированными в единый фенон, хорошо обособленный от остальных видов рода. Внутри него минимальный уровень различий свойствен рыбам Антарктического полуострова и Ю. Шетландских о-вов, что подтверждается результатами расчетов фенетических дистанций с применением всех алгоритмов кластеризации, используемых в программе «SYSTAT». Скорее всего, обе эти выборки принадлежат к единой «южной» популяции зеленой нототении, наибольшая численность которой в пределах видового ареала обеспечивает поддержание внутри нее достаточно высокой степени панмиксии. Высокая численность этой популяции подтверждается как данными промысловой статистики, так и подсчетами численности личинок (Worner, James, 1981). Весьма вероятно, что пойманные у берегов Антарктического полуострова рыбы представляют часть нагульного стада, мигрирующего на юг в летнее (для южного полушария) время.*
Результаты расчетов фенетических дистанций по большинству (5 из 6) алгоритмов кластеризации показывают, что две южные выборки G. gibberifrons (Антарктического полуострова и Ю. Шетландских островов) более сходны с рыбами ближайших к ним Ю. Оркнейских островов, чем с о. Ю. Георгия (рис. 1, А, Б). Это подтверждает предыдущие оценки общего сходства зеленой нототении из этих 4 районов Западной Антарктики, которые были получены на меньших по объему и числу признаков материалах (Балушкин, 1991). Связь общего фенотипического сходства рыб и географического положения районов их обитания указывает на то, что степень морфологической обособленности определяется главным образом межпопуляционным обменом, при котором «перемешивание» рыб реже и труднее осуществляется между более удаленными друг от друга территориями.
_________________
* Через пролив Брансфилд и далее на северо-восток вдоль вторичной фронтальной зоны проходит зоогеографическая граница, разделяющая две исторически сложившихся ихтиофауны: континентальную циркумантарктическую и островную западноантаркти-ческую (фауну рыб островов моря Скотия) (Balushkin, 2000). Географическая близость Ю. Шетландских островов к материку обусловливает сгущение ареалов видов обеих фаун во многом за счет сезонных миграций рыб, осуществлению которых благоприятствуют изменения температурного, гидрологического и ледового режимов в данном районе. Это обстоятельство, на наш взгляд, определяет полосной характер (термин Б.А. Кузнецова, 1936) синператы, охватывающей акватории от северной оконечности Антарктического полуострова до Южных Шетландских островов. Выделение этой пограничной полосы в качестве особого зоогеографического выдела в ранге провинции или округа (Андрияшев, 1986; Смирнов, 1990, 1992) нам представляется не целесообразным.
Летом у берегов Антарктического полуострова вместе с зеленой нототенией встречаются и другие западноантарктические виды: Nototheniops nybelini (Balushkin), Lindbergichthys nudifrons (Lonnberg), Parachaenichthys charcoti (Vaillant), где они интенсивно питаются крилем (по данным А.В. Балушкина, полученным в экспедиции на НИС «Гижига», 1984–1985 гг.).
Среди видов Gobionotothen устойчивые отношения общего сходства (по всем алгоритмам кластеризации) характерны для нототении-губошлепа G. angustifrons и марионской бычковидной нототении G. marionensis.
Некоторые авторы (Gon, Klages, 1988; DeWitt et al., 1990) считают эти виды конспецифичными, но без достаточных на то оснований (Балушкин, 1991). Они аллопатричны и хорошо отличимы друг от друга числом лучей в анальном плавнике и особенностями очешуения головы, а высокую степень изоляции друг от друга этих мелких литоральных видов обеспечивают огромные океанические расстояния (около 60 градусов по широте) и серьезный гидрологический барьер (зона антарктической конвергенции).
В большинстве случаев топология сходств видов Gobionotothen имеет вид G. marionensis, G. angustifrons) G. acuta)) G. barsukovi, G. gibberifrons (рис.1), и лишь в одном (рис. 2) – G. marionensis, G. angustifrons) G. acuta, G. barsukovi)) G. gibberifrons. Неопределенность положения G. barsukovi на дендрограммах, полученных разными методами кластеризации, связана с тем, что многие морфометрические признаки имеют у этого вида промежуточные значения между, с одной стороны, мелкими видами рода, объединяемыми в группу видов «marionensis» (G. marionensis, G. angustifrons и G. acuta), с другой – G. gibberifrons (Балушкин, 1991, табл. 1–3). С последним видом G. barsukovi разделяет ряд четких диагностических признаков (большое число чешуй в боковых линиях, лучей в анальном плавнике, позвонков, более сильное очешуение головы и передней части туловища, более крупные размеры взрослых рыб), что давало веские основания выделять их в качестве отдельной группы видов «gibberifrons». Вместе с тем наличие определенных сходств между G. barsukovi и смежно-симпатрическим, обитающим на меньших глубинах мелким видом G. acuta (высокий, треугольной формы колючий спинной плавник, число чешуй в дорсальной боковой линии) указывало на возможный парафилетический характер видовых групп рода Gobionotothen (Балушкин, 1991). Как видно из табл. 1, G. barsukovi и G. acuta также отличаются от других видов меньшей длиной рыла.
В Западной Антарктике существует сходная смежно-симпатрическая пара видов, которая включает мелкий литоральный вид (G. angustifrons) и более крупный глубоководный вид – G. gibberifrons. Морфологическая специфика этой пары заключается в том, что ее видам (при сравнении с их аналогами из «кергеленской» пары) свойственны согласованные изменения некоторых признаков. Так, у G. gibberifrons число жаберных тычинок внешнего ряда в среднем в 1.1 раза меньше, чем у G. barsukovi, и, соответственно, у G. angustifrons – в 1.1 раза меньше, чем у G. acuta. У G. gibberifrons высота головы (Ho) в 1.2 раза больше, чем у G. barsukovi, и у G. angustifrons – в 1.1 раза больше, чем у G. acuta. Показательно при этом, что G. marionensis – морфологически близкий к G. angustifrons (см. выше) и, вероятно, сравнительно недавно обособившийся от него вид сходным образом отличен в приведенных выше признаках от G. acuta, как и G. angustifrons. Отмеченные особенности позволяют предполагать происхождение обеих пар видов от единой предковой формы, развитие которой в определенном направлении приводит к независимому появлению тождественных форм, не имеющих прямой генетической связи друг с другом. Такой способ видообразования находит свое объяснение в рамках теории номогенеза Л.С. Берга, обобщающей различные проявления полифилетизма, в том числе политопное образование форм (=изокинетическое состояние форм А.П. Семенова-Тян-Шанского) и «повторное видообразование» Кокена, или итеративная эволюция (Берг, 1977; Балушкин, 2002).
Предполагаемый характер кладогенеза бычковидных нототений иллюстрирует схема на рис. 3, показывающая независимое образование сходных форм (G. gibberifrons и G. barsukovi, с одной стороны, и G. angustifrons и G. acuta – с другой) в ходе эволюции предковой формы. Пути ее расселения в Южном океане могли быть различными: она могла проникнуть из Западной Антарктиды на Кергеленское плато либо океаническим путем (при посредстве келпов – островов бурых водорослей, дрейфующих в Южном океане под влиянием течения западных ветров), либо могла расселяться вдоль материка и проникнуть на Кергеленское плато в один из периодов регрессии моря (в частности, во время крупной регрессии на границе плиоцена-плейстоцена) (Балушкин, 1991; Balushkin, 2000). В последнем случае исчезновение предковой формы у берегов Антарктиды, приведшее к дизъюнкции ранее обширного единого ареала, могло произойти во время одного из плейстоценовых оледенений Восточной Антарктиды, вызвавшего катастрофические последствия для прибрежной континен-тальной фауны в целом. Общий уровень морфологических различий западноантарктических видов друг от друга значительно выше, чем у видов кергеленской пары. Этот факт может косвенно подтверждать направленность расселения предковой формы с запада на восток, а также то, что в отношении начальных фаз дивергенции процесс видообразования шел не только политопно, но и гетерохронно. Акты видообразования были разобщены, по меньшей мере, на то время, которое потребовалось предковой форме для проникновения на Кергеленское плато.
В ходе становления эндемичной антарктической ихтиофауны кладогенез бычковидных нототений является не единственным примером итеративной эволюции. На наш взгляд, такой способ повторного видообразования (или, по Семенову-Тян-Шанскому, изокинетического превращения) был характерен также для антарктических рогаток рода Harpagifer (Балушкин, 2002). Заселяя отдаленные от материка острова Южного океана, находящиеся в зоне циркумантарктического течения, рогатки неоднократно дивергировали на две морфологически и батопатически сходные формы: “литоральную” и “глубинную” (или “рогатую”, по выражению Андрияшева и Токарева, 1958). Первая форма представлена видами H. marionensis (Nybelin, 1947) у о. Марион, H. kerguelensis (Nybelin, 1947) у о-вов Кергелен, H. macquariensis Prirodina, 2000 у о. Маккуори, вторая – H. spinosus Hureau, Louis, Tomo et Ozouf, 1980) у о-вов Крозе, H. nybelini Prirodina, 2002 у о-вов Кергелен и H. andriashevi Prirodina, 2000 у о. Маккуори (Природина, 2000, 2002). Случаи повторного видообразования можно предполагать и у антарктических щекорогов из преимущественно периантарктического рода Bovichtus (сем. Bovichtidae), виды которого известны у Ю. Америки (B. chilensis Regan, 1913), юго-восточной Австралии и Тасмании (B. angustifrons Regan, 1913), Новой Зеландии и близлежащих островов (B. variegatus Richardson, 1844), Антарктического полуострова (B. elongatus Hureau et Tomo, 1977), а также у отдаленных островов в южных частях Атлантического, Индийского и Тихого океанов: Тристан-да-Кунья и Гоф (B. diacanthus (Carmichael, 1818), Сен-Поль и Амстердам (B. veneris Sauvage, 1879) и у островов Новозеландского плато (B. psychrolutes Gunther, 1860). Взрослые экземпляры этих видов живут на литорали, среди камней, прибрежных скал и рифов в приливно-отливной зоне на глубинах не более 30 м, и лишь один вид (B. oculus Hardy, 1988) был сравнительно недавно описан с глубин 800–900 м от о. Кемпбелл. В данном месте глубоководный B. oculus составляет смежно-симпатрическую пару литоральному виду B. psychrolutes, который, в отличие от другого новозеландского щекорога – B. variegatus, населяет наиболее окраинные острова юга и юго-востока Новозеландского плато. B. oculus и B. psychrolutes близки морфологически, хорошо отличаясь от других австралазийских видов наличием пятна чешуй под грудным плавником и рядом меристических признаков (Hardy, 1988). Происходили ли сходные процессы в других районах обитания антарктических щекорогов, покажут будущие исследования.
Рис. 1. Дендрограммы различий видов Gobionotothen и 4 выборок G. gibberifrons по 24 морфологическим признакам. Алгоритмы кластеризации: А - Average Linkage Method, Б – Complete Linkage Method (farthest neighbor). Обозначения районов: А.П. – Антарктический п-ов (юго-восточная часть прол. Брансфилд), Ю.Г. – о. Южная Георгия, Ю.О. – Южные
Оркнейские о-ва, Ю.Ш. – Южные Шетландские о-ва.

Рис. 2. Дендрограмма различий видов Gobionotothen и 4 выборок G. gibberifrons по 24 морфологическим признакам. Алгоритм кластеризации: Ward Minimum Variance Method. Обозначения как на рис. 1.

Рис. 3. Схема, иллюстрирующая эффект повторного видообразования (итеративной эволюции) у бычковидных нототений рода Gobionotothen (объяснение в тексте).
Таблица 1. Морфометрические признаки зеленых нототений рода Gobionotothen
|
Измерения |
Признак |
G. gibberifrons Антарктич. п-ов |
G. gibberifrons Ю.Шетланд. о-ва |
G. gibberifrons Ю.Оркнейские о-ва |
G. gibberifrons о. Ю.Георгия |
G. barsukovi |
G. angustifrons |
G. marionensis |
G. acuta |
|||||||
|
x |
lim |
x |
lim |
x |
lim |
x |
lim |
x |
lim |
x |
lim |
x |
lim |
x |
||
|
В % SL |
aD1 |
30.9 |
29.4-32.5 |
31.4 |
29.2-33.4 |
30.9 |
28.9-32.5 |
31.1 |
29.6-32.2 |
30.9 |
28.2-33.1 |
33.9 |
31.1-37 |
35.1 |
34-36.1 |
35 |
|
aD2 |
42.1 |
39.8-45 |
42.5 |
39.5-44.6 |
42.2 |
37.3-44.8 |
41.7 |
40.3-43.9 |
42 |
38.8-47.2 |
45.3 |
36.7-48.6 |
48.4 |
46.5-49.7 |
45.7 |
|
|
aA1 |
45.4 |
42-47.7 |
45.2 |
43.4-48 |
46 |
43-50 |
42.7 |
40.9-44.9 |
43.4 |
39.8-46.1 |
48.2 |
45.1-53.8 |
52.9 |
50.6-55 |
49 |
|
|
LP |
25.4 |
23.1-27 |
23.7 |
19.4-26.1 |
25.2 |
23.1-27 |
22.3 |
21.2-23.9 |
23.4 |
18.4-24.4 |
23 |
20.6-27.1 |
23.3 |
22.3-24.8 |
26.1 |
|
|
LV |
18 |
15.5-19.4 |
17.1 |
16.6-19.4 |
17.1 |
15.5-20.7 |
16.5 |
15-18.2 |
17 |
14.9-20 |
19.2 |
15.3-22.8 |
19.8 |
17.8-21.3 |
20.1 |
|
|
Lcp |
6.8 |
6.2-7.3 |
7 |
6-8 |
7.2 |
6.2-9.4 |
7.1 |
6.4-8.7 |
5.5 |
4.6-6.3 |
5.5 |
4-6.6 |
4.8 |
4.3-5.6 |
6.6 |
|
|
Hcp |
5.9 |
5.5-6.3 |
6.1 |
5.5-6.5 |
5.6 |
4.7-6.4 |
6.1 |
5.5-6.8 |
6.1 |
5.5-6.9 |
5.7 |
5.2-6.6 |
6.3 |
6.1-6.4 |
7.5 |
|
|
Lc1 |
26.6 |
25.3-27.9 |
26 |
24.8-26.7 |
26 |
23.9-27.6 |
26.2 |
24.9-27.2 |
25.4 |
23.3-28.7 |
29.4 |
26.4-31.4 |
28.5 |
28.2-28.8 |
29.7 |
|
|
В % Lc1 |
Wc |
67.8 |
56.6-75.9 |
68.1 |
57.2-78 |
66.4 |
54.7-71.8 |
66.1 |
62-70.4 |
63 |
50-71 |
60.1 |
53.4-71.7 |
64.7 |
60-68 |
59.5 |
|
Ho |
48.1 |
45.7-51.1 |
49.1 |
43.8-53.3 |
46.4 |
41.1-49.4 |
46.1 |
43.3-48.4 |
40.8 |
38.2-44.9 |
43.4 |
38.6-51 |
42.2 |
41.4-43.4 |
39.9 |
|
|
aO |
31 |
29.1-33.9 |
30.7 |
24.9-33.5 |
29.2 |
25.8-32.1 |
33.5 |
31-35.7 |
28.4 |
26-30.8 |
31.4 |
28.3-34.7 |
29.8 |
29-30.9 |
27.8 |
|
|
O |
24.1 |
21.6-26.9 |
26.2 |
23.1-29.8 |
27.8 |
24.3-32.8 |
26.7 |
23.9-29.7 |
27.5 |
25.1-29.8 |
26.8 |
20.4-31.5 |
25.2 |
24.4-26.3 |
30.4 |
|
|
Lmx |
32.3 |
30.6-33.9 |
32.2 |
29-35.1 |
32.9 |
30-34.9 |
31.4 |
29.3-34.3 |
32.9 |
28.1-35.9 |
33.9 |
26.1-37.7 |
34.3 |
33.3-36.1 |
32.3 |
|
|
Lmd |
35 |
32.7-37.5 |
35.2 |
31.5-37.9 |
36.4 |
33.9-40.7 |
34.8 |
31.7-37.8 |
37.1 |
32.8-39.9 |
41 |
37.7-44.1 |
37.5 |
35.4-39.6 |
39.9 |
|
|
iO |
7.5 |
6.6-8.5 |
8.2 |
6.5-10.2 |
6.7 |
5.5-8 |
7.6 |
6.8-8.3 |
6.5 |
5-7.9 |
5.9 |
3.6-9 |
7.4 |
6.7-8 |
7 |
|
|
Подсчеты |
D1 |
7.14 |
6-8 |
7.11 |
6-8 |
7.01 |
6-8 |
7.13 |
6-8 |
6 |
5-7 |
5.9 |
4-7 |
6.5 |
6-7 |
6 |
|
D2 |
32.17 |
30-34 |
32.1 |
31-34 |
32.26 |
31-34 |
32.17 |
31-34 |
31.1 |
30-33 |
29.7 |
28-31 |
29 |
28-30 |
30 |
|
|
A |
32.07 |
31-33 |
32.06 |
30-34 |
32.13 |
31-33 |
31.9 |
31-33 |
32.2 |
31-34 |
30 |
29-31 |
28.2 |
27-29 |
30 |
|
|
P |
20.95 |
20-22 |
21.14 |
20-23 |
21.34 |
20-23 |
20.9 |
19-23 |
20.4 |
19-21 |
19.8 |
19-21 |
19.8 |
19-21 |
20 |
|
|
ULL |
41.3 |
39-45 |
42.7 |
39-46 |
42.6 |
40-45 |
41.1 |
37-43 |
39.8 |
36-44 |
31.1 |
27-35 |
32 |
25-35 |
36.5 |
|
|
MLL |
35.6 |
31-41 |
35.6 |
31-44 |
37.9 |
31-42 |
34.7 |
29-38 |
28.2 |
22-33 |
17.1 |
13-26 |
15.8 |
14-19 |
16.5 |
|
|
Sp.br.ant. |
17 |
16-19 |
17.3 |
15-19 |
16.8 |
13-20 |
17.9 |
17-19 |
18.7 |
17-20 |
17.5 |
15-20 |
17.5 |
17-18 |
19 |
|
|
Sp.br.post. |
11.8 |
11-13 |
12.4 |
10-13 |
12 |
11-14 |
12.1 |
11-14 |
11.3 |
11-13 |
11 |
10-12 |
11.5 |
11-12 |
11 |
|
|
vert. |
49.6 |
48-51 |
49.7 |
48-51 |
49.8 |
48-51 |
49.6 |
48-52 |
47.5 |
47-49 |
45.3 |
44-46 |
44.2 |
44-45 |
46 |
|
К МОРФОЛОГИЧЕСКОЙ ХАРАКТЕРИСТИКЕ МРАМОРНОЙ НОТОТЕНИИ NOTOTHENIA ROSSII RICHARDSON, 1844 (NOTOTHENIIDAE) ОСТРОВОВ КЕРГЕЛЕН
Один из важнейших промысловых видов Южного океана - мраморная нототения обитает у всех островов моря Скотия (Южная Георгия, Южные Сандвичевы, Южные Оркнейские и Южные Шетландские острова), в проливе Брансфилд, в северо-восточной части моря Беллинсгаузена (до Земли Александра I) и на севере Антарктического полуострова. В индоокеанском секторе Антарктики вид известен у островов Принс-Эдуард, Крозе, Кергелен, Х¨рд, Маккуори, на банках Обь, Лена и Скиф (DeWitt et al., 1990; наши данные). Этому виду посвящено большое число работ по биологии и промыслу (последний обзор см.: Шуст, 1998) и, в меньшей мере, – изучению его морфологии. Последнее обстоятельство в особенности относится к популяции вида из районов индоокеанского сектора Южного океана. Краткие описания внешнего строения рыб из этого района содержатся в ряде ранних таксономических описаний (Richardson, 1844; Norman, 1937; Nybelin, 1947; Hureau, 1970 и др.), однако только в статье П.Б. Танкевича (1986) была дана основанная на массовых материалах развернутая морфометрическая характеристика кергеленской мраморной нототении.
Цель настоящей работы – дать описание и анализ изменчивости головных каналов сейсмосенсорной системы и провести сравнение литературных (Танкевич, 1986) и собственных данных по морфометрии мраморной нототении из индоокеанского сектора Южного океана.
Система головных каналов боковой линии у мраморной нототении Notothenia rossii Richardson
Супраорбитальный (надглазничный) канал (canalis supraorbitalis, CSO) проходит в nasale и frontale, соединяясь над глазом с одноименным каналом противоположной стороны, сзади – с подглазничным и височным каналами. У всех исследованных экземпляров передняя часть канала до корональной коммиссуры состоит из 3 сегментов и имеет 3 основные поры (pso1-pso3), открывающиеся на канале латерально (к глазу). Посткорональная часть канала содержит 2 сегмента и 1 пору (pso5), открывающуюся между ними к центру тела (от глаза). Последний сегмент канала (5-й по счету) имеет, кроме того, общую основную пору с первым сегментом темпорального и последним сегментом (=dermosphenoticum) инфраорбитального каналов. По принятой у нототениевых терминологии (Якубовски, 1971; Балушкин, 1984) эта пора считается 1-й порой темпорального канала (pt1). Индивидуальная изменчивость в числе пор была обнаружена нами только в посткорональной (заглазничной) части канала: у 3 экз. слева и у 1 экз. справа вместо одной основной поры (pso5) появляются две межсегментные дополнительные поры (pso5a и pso5b). Ранее такое увеличение пор (причем только с одной стороны тела) было отмечено не только у мраморной нототении, но и одного из близких к ней видов – антарктической нототении N. coriiceps Richardson, 1844 (Балушкин, 1984). У нототениевых закрепление этой индивидуальной вариации в норму происходит у патагонских нототений Patagonotothen Balushkin, у которых на этом участке канала формируется также дополнительный канальный невромаст (т.е. создается новый сегмент канала).
Корональная коммиссура (commissura coronalis, CC) соединяет надглазничные каналы обеих сторон головы. Она не имеет канальных невромастов и открывается одной порой (pcor), которая фактически является 4-й порой супраорбитального канала (pso4), общей для аналогичных сегментов CSO обеих сторон тела. Корональная коммиссура присутствует у всех исследованных экземпляров Notothenia rossii.
Инфраорбитальный (подглазничный) канал (canalis infraorbitalis, CIO) проходит в слезной и постлакримальных костях. Последним сегментом канала является dermosphenoticum. Канал соединяется позади глаза с супраорбитальным и темпоральным каналами. Общая пора этих 3-х каналов (pt1) заметно смещена назад от заднего края глаза (как и у других видов Notothenia Richardson). Число пор в CIO наиболее изменчиво, варьируя от 7 до 11 (табл. 2). Наиболее часто встречаются каналы с общим числом пор 8–9, из которых 5 пор расположены на lacrimale. Такое строение наблюдалось в 77.5% случаев (табл. 3), что практически совпадает с прежними подсчетами (77.4% по данным Балушкина, 1984, табл. 6).
Таблица 2. Число пор в инфраорбитальном канале у Notothenia rossii Richardson
|
Пор в lacrimale |
Всего пор в канале |
Число экз. |
||
|
слева |
справа |
слева |
справа |
|
|
4 |
4 |
8 |
7 |
1 |
|
5 |
4 |
9 |
7 |
1 |
|
5 |
5 |
8 |
8 |
7 |
|
5 |
5 |
8 |
9 |
1 |
|
5 |
5 |
9 |
8 |
4 |
|
5 |
5 |
9 |
9 |
2 |
|
5 |
5 |
9 |
10 |
1 |
|
5 |
5 |
10 |
10 |
1 |
|
5 |
5 |
11 |
11 |
1 |
|
6 |
6 |
9 |
9 |
1 |
Темпоральный (височный) канал (canalis temporalis, CT) проходит через frontale, pteroticum, tabulare supratemporale и posttemporale. Через сегмент в supracleithrum он переходит в туловищный канал. Типичным для всех родов подсемейства Nototheniinae является 5 пор в темпоральном канале (сегмент, проходящий через supracleithrum, относится не к височному, а к туловищному каналу: по Якубовски, 1971). Первые 4 поры открываются латерально, а 5-я – медиально. У исследованных нами экземпляров изменчивости в числе пор не обнаружено – все они имели по 5 пор с обеих сторон головы. Примеры индивидуальных вариаций в строении CT, полученные при изучении других коллекций мраморной нототении, приведены ранее Балушкиным (1984).
Таблица 3. Различные сочетания числа пор в слезной кости и в инфраорбитальном канале в целом и частота их встречаемости (в %) у Notothenia rossii Richardson
(у 20 экз. с обеих сторон тела)
Пор в lacrimale |
Всего пор в канале |
Встречаемость,% |
4 |
7 |
5 |
4 |
8 |
2.5 |
5 |
8 |
50 |
5 |
9 |
27.5 |
5 |
10 |
7.5 |
5 |
11 |
5 |
6 |
9 |
5 |
Супратемпоральная (надвисочная) коммиссура (commissura supratemporalis, CST), соединяющая левый и правый темпоральные каналы, проходит с каждой стороны головы в двух табличных косточках (tabulare parietale и tabulare supraoccipitale). Notothenia rossii, как и большинство видов подсемейства, имеет 3 основные поры в CST. Хотя изменений в числе пор надвисочной коммиссуры на данном материале нами обнаружено не было, у мраморной нототении число пор здесь может изредка возрастать до 4-х из-за появления дополнительной сегментной поры на одном из сенсов справа или слева (Балушкин, 1984).
Предкрышечно-нижнечелюстной, или преоперкуломандибулярный канал (canalis praeoperculo–mandibularis, CPM) проходит в dentale, anguloarticulare и praeoperculum. Число основных пор у видов подсемейства нототениевых, не имеющих связи CPM с CT, строго постоянно (10 пор) (Балушкин, 1984). Изменчивости в строении этого канала у исследуемых здесь рыб не обнаружено. Ранее у одного экземпляра этого вида с левой стороны головы был описан случай соединения CPM с CT, сопровождавшийся соответствующей потерей одной поры в канале (Балушкин, 1984), что следует рассматривать как индивидуальную реверсию предкового (плезиоморфного) состояния.
Морфометрическая характеристика мраморной нототении
Наши данные по морфометрии мраморной нототении о-вов Кергелен, включающие 9 меристических и 24 пластических признака, представлены в табл. 4. Их сравнение с данными Танкевича (1986), основанными на выборках рыб от о-вов Кергелен и банки Лена, дано по 22 сопоставимым признакам с использованием коэффициента различия CD Майра (табл. 5–7).
Проведенные нами расчеты показали, что по всем использованным для сравнения признакам общее неперекрывание кривых распределения было ниже 82%, т.е. коэффициент различия никогда не превышал 0.91, а в большинстве случаев он был значительно меньше этого значения (табл. 5 и 6). Такие незначительные отличия сравниваемых выборок свидетельствуют в пользу вывода Танкевича (1986) о том, что у о-вов Кергелен на банке Лена обитает единая, морфологически однородная популяция мраморной нототении.
Поскольку общие размеры рыб в нашей коллекции были в среднем ниже, чем у П.Б. Танкевича, то часть статистически достоверных отличий, касающихся пропорций тела, может быть объяснена возрастной изменчивостью. Действительно, при сравнении нашей выборки лишь с той частью выборки Танкевича, размеры рыб в которой не превышали 400 мм SL, значения коэффициента различия Майра уменьшаются для большинства пластических признаков (ср. табл. 5 и 7). Сравнивая 2 размерные группы (293–399 мм SL и 449–658 мм SL), Танкевич установил статистически достоверные изменения некоторых пропорций тела: у мраморной нототении с возрастом уменьшаются высота спинных плавников (hD1, hD2), длина брюшного плавника (LV), наименьшая высота тела (=высота хвостового стебля, hcp) и диаметр глаза (O). Кроме того, у более крупных рыб уменьшается межглазничное расстояние (iO кожн.), увеличивается антеанальное расстояние (aA1). В соответствие с этими изменениями могут быть поставлены и различия между нашей выборкой и обеими выборками Танкевича в таких признаках, как hD1, hD2, hcp, iO кожн. и aA1.
Танкевич (1986) справедливо предполагает возможность пассивного переноса (в струях Антарктического циркумполярного течения) оплодотворенной икры и молоди мраморной нототении с акватории банки Лена на шельф о-вов Кергелен, что и обусловливает морфологическое сходство рыб исследованных районов. К этому можно только добавить, что наличие стадии пелагических мальков у этого вида значительно повышает шансы успешной эмиграции «ленских» рыб к островам Кергеленского архипелага.
Таблица 4. Морфометрические признаки мраморной нототении (о-ва Кергелен, n =20)
|
Признак |
lim |
x±mx |
σ |
|
SL, мм |
214.2-378.3 |
308.11±11.55 |
30.34 |
|
ID |
5-8 |
5.85±0.20 |
0.88 |
|
IID |
32-35 |
33.65±0.21 |
0.93 |
|
A |
26-29 |
27.50±0.19 |
0.83 |
|
P |
21-23 |
22.10±0.08 |
0.5 |
|
ULL |
43-55 |
49.00±0.51 |
3.17 |
|
MLL |
12-23 |
16.35±0.34 |
2.12 |
|
Sp.br.ant. |
13-22 |
19.25±0.50 |
2.17 |
|
Sp.br.post. |
10-15 |
13.00±0.30 |
1.30 |
|
Sq |
51-59 |
54.10±0.37 |
2.33 |
|
В % SL |
|||
|
H |
19.53-26.36 |
22.39±0.41 |
1.79 |
|
aD1 |
26.65-31-68 |
30.06±0.19 |
0.84 |
|
HD1 |
4.69-6.78 |
5.59±0.14 |
0.60 |
|
aD2 |
37.48-44.69 |
41.81±0.36 |
1.57 |
|
HD2 |
8.21-12.42 |
10.38±0.30 |
1.29 |
|
aA11 |
46.28-56.96 |
50.95±1.96 |
8.53 |
|
aA2 |
27.13-57.36 |
51.09±1.95 |
8.49 |
|
ABr |
14.80-24.88 |
17.60±0.69 |
3.02 |
|
LP |
17.52-22.80 |
20.15±0.37 |
1.61 |
|
LV |
9.10-16.78 |
13.41±0.72 |
2.16 |
|
LVA |
22.29-30.48 |
26.20±0.45 |
1.98 |
|
Lср. |
9.57-12.73 |
11.36±0.19 |
0.82 |
|
Hср. |
7.57-10.11 |
8.97±0.13 |
0.56 |
|
Lс1 |
24.53-30.71 |
28.79±0.30 |
1.29 |
|
Lc2 |
12.55-20.18 |
18.59±0.40 |
1.72 |
|
В % длины головы (Lc1) |
|||
|
Wс |
63.26-88.32 |
76.63±1.73 |
7.54 |
|
Hc |
62.97-81.39 |
70.40±1.22 |
5.33 |
|
Ho |
48.74-65.75 |
54.27±1.10 |
4.81 |
|
AO |
26.45-34.23 |
28.96±0.44 |
1.92 |
|
O |
16.62-21.77 |
18.88±0.32 |
1.40 |
|
Lmx |
36.96-44.33 |
41.99±0.41 |
1.81 |
|
Lmd |
47.40-52.52 |
49.89±0.34 |
1.48 |
|
iO (кожное) |
29.83-35.62 |
31.78±0.37 |
1.62 |
|
iO (костное) |
25.81-32.03 |
28.33±0.40 |
1.75 |
Таблица 5. Морфометрические признаки и значения коэффициента различия (CD) у двух выборок мраморной нототении N. rossii о-вов Кергелен
|
Признак |
Наши данные (n=20) |
Данные Танкевича (1986) (n=73) |
|||||
|
|
lim |
x±mx |
σ |
lim |
x±mx |
σ |
CD |
|
SL, мм |
214.62-378.3 |
308 ±11.55 |
50.34 |
293-658 |
413.18±8.65 |
38.24 |
1.19 |
|
ID |
5-8 |
5,85±0.20 |
0.88 |
5-7 |
5.78±0.08 |
0.63 |
0.05 |
|
IID |
32-35 |
33.65±0.21 |
0.93 |
32-37 |
33.75±0.10 |
0.85 |
0.06 |
|
A |
26-29 |
27.50±0.19 |
0.83 |
24-29 |
26.65±0.10 |
0.81 |
0.52 |
|
P |
21-23 |
22.10±0.08 |
0.50 |
20-23 |
21.15±0.09 |
0.78 |
0.74 |
|
ULL |
43-55 |
49.45±0.51 |
3.17 |
39-57 |
48.96±0.49 |
4.13 |
0.12 |
|
MLL |
12-22 |
16.35±0.34 |
2.12 |
11-22 |
16.78±0.27 |
2.25 |
0.10 |
|
Sp.br.ant. |
13-22 |
19.25±0.50 |
2.17 |
16-21 |
18.97±0.14 |
1.20 |
0.08 |
|
В % SL |
|||||||
|
H |
19.53-26.36 |
22.39±0.41 |
1.76 |
19.4-28 |
23.47±0.23 |
1.93 |
0.29 |
|
aD1 |
26.65-31.68 |
30.06±0.19 |
0.84 |
27.9-35 |
29.75±0.16 |
1.34 |
0.14 |
|
HD1 |
4.69-6.78 |
5.59±0.14 |
0.60 |
3.8 –5.9 |
4.91±0.06 |
0.50 |
0.62 |
|
HD2 |
8.21-12.42 |
10.38±0.30 |
1.29 |
7.5-11 |
9.28±0.09 |
0.75 |
0.54 |
|
aA1 |
46.28-56.96 |
50.95±1.96 |
8.53 |
47.9-55.4 |
52.79±0.29 |
2.43 |
0.17 |
|
LP |
17.52-22.80 |
20.51±0.37 |
1.61 |
16.8-22.7 |
20.34±0.13 |
1.13 |
0.06 |
|
LV |
9.10-16.78 |
13.41±0.72 |
2.16 |
9.80-16.7 |
14.80±0.16 |
1.38 |
0.39 |
|
Hср |
7.57-10.11 |
8.97±0.13 |
0.56 |
6.6-9.7 |
8.40±0.08 |
0.64 |
0.48 |
|
Lc1 |
24.53-30.71 |
28.79±0.30 |
1.28 |
25.3-30.6 |
28.63±0.13 |
1.12 |
0.07 |
|
В % длины головы (Lc1) |
|||||||
|
Ho |
26.45-34.23 |
26.96±0.44 |
1.92 |
27.2 -35.3 |
30.98±0.20 |
1.72 |
0.39 |
|
O |
16.62-21.77 |
18.88±0.32 |
1.40 |
16.4-23 |
19.84±0.16 |
1.35 |
0.35 |
|
Lmx |
36.96-44.33 |
41.99±0.41 |
1.81 |
38.8-45.1 |
42.16±0.18 |
1.49 |
0.05 |
|
Lmd |
47.40-52.52 |
49.89±0.34 |
1.48 |
43.9-54.3 |
48.27±0.26 |
2.23 |
0.44 |
|
IO кожн |
29.83-35.62 |
31.78±0.37 |
1.62 |
26.7-33.2 |
30.19±0.17 |
1.41 |
0.52 |
Таблица 6. Морфометрические признаки и значения коэффициента различия (CD) мраморной нототении N. rossii о-вов Кергелен и банки Лена
|
Признак |
Наши данные (n=20) |
Данные Танкевича (1986) (n=73) |
|||||
|
о. Кергелен |
банка Лена |
||||||
|
|
lim |
x±mx |
σ |
lim |
x±mx |
σ |
CD |
|
SL, мм |
214.2-378.3 |
308.11±11.55 |
50.34 |
325.0-610.0 |
424.86±6.79 |
37.27 |
1.33 |
|
ID |
5-8 |
5.85±0.20 |
0.88 |
5-7 |
5.65±0.10 |
0.84 |
0.1 |
|
IID |
32-35 |
33.65±0.21 |
0.93 |
32-37 |
33.81±0.12 |
0.96 |
0.08 |
|
A |
26-29 |
27.5±0.19 |
0.83 |
25-29 |
26.65±0.11 |
0.87 |
0.50 |
|
P |
21-23 |
22.10±0.08 |
0.50 |
20-22 |
20.90±0.10 |
0.84 |
0.90 |
|
ULL |
43-55 |
49.45±0.51 |
3.17 |
42-57 |
48.50±0.51 |
4.21 |
0.13 |
|
MLL |
12-22 |
16.35±0.34 |
2.12 |
11-21 |
16.79±0.20 |
1.69 |
0.12 |
|
Sp.br.ant. |
13-22 |
19.25±0.50 |
2.17 |
16-21 |
18.61±0.14 |
1.15 |
0.19 |
|
В % SL |
|||||||
|
H |
19.53-26.36 |
22.39±0.41 |
1.79 |
17.1-29.1 |
23.41±0.31 |
2.55 |
0.24 |
|
aD1 |
26.63-31.68 |
30.06±0.19 |
0.84 |
26.4-33.2 |
29.9±0.15 |
1.22 |
0.08 |
|
HD1 |
4.69-6.78 |
5.59±0.14 |
0.60 |
3.0-6.6 |
4.67±0.08 |
0.69 |
0.71 |
|
HD2 |
8.21-12.42 |
10.38±0.30 |
1.29 |
7.5-11.0 |
8.96±0.11 |
0.87 |
0.66 |
|
aA1 |
46.28-56.96 |
50.95±1.96 |
8.53 |
47.9-55.4 |
51.78±0.43 |
3.55 |
0.07 |
|
LP |
17.52-22.80 |
20.51±0.37 |
1.61 |
16.8-22.7 |
20.26±0.15 |
1.23 |
0.09 |
|
LV |
9.10-16.78 |
13.41±0.72 |
2.16 |
9.80-16.7 |
14.41±0.17 |
1.42 |
0.28 |
|
Hср |
7.57-10.11 |
8.97±0.13 |
0.56 |
6.6-9.7 |
8.29±0.08 |
0.68 |
0.55 |
|
Lc1 |
24.53-30.71 |
28.79±0.30 |
1.28 |
25.3-30.6 |
28.33±0.14 |
1.19 |
0.19 |
Таблица 7. Морфометрические признаки и значения коэффициента различия (CD) у двух выборок мраморной нототении N. rossii о-вов Кергелен размерной группы до 399 мм SL
|
Признак |
Наши данные (n=20) |
Данные Танкевича (1986) (n=34) |
|||||
|
|
lim |
x ± mx |
σ |
lim |
x± mx |
σ |
CD |
|
SL |
214.2-378.3 |
308.11± 11.55 |
50.34 |
293.0-399.0 |
356.94± 4.82 |
28.12 |
0.62 |
|
В % SL |
|||||||
|
H |
19.53-26.36 |
22.39±0.41 |
1.76 |
20.4-28.0 |
23.5± 0.34 |
2.01 |
0.29 |
|
aD1 |
26.65-31.68 |
30.06±0.19 |
0.84 |
28.92-35.0 |
28.92±0.21 |
1.21 |
0.56 |
|
HD1 |
4.69-6.78 |
5.59±0.14 |
0.60 |
4.7-5.9 |
5.21±0.07 |
0.38 |
0.32 |
|
HD2 |
8.21-12.42 |
10.98±0.30 |
1.29 |
8.1-11.1 |
9.61±0.11 |
0.66 |
0.39 |
|
aA1 |
46.28-56.96 |
50.95±1.96 |
8.53 |
48.8-59.5 |
52.42±0.36 |
2.08 |
0.14 |
|
LP |
17.52-22.80 |
20.51±0.37 |
1.61 |
17.2-23.3 |
20.21±0.19 |
1.09 |
0.11 |
|
LV |
9.10-16.78 |
13.41±0.72 |
2.16 |
13.9-18.6 |
15.68±0.16 |
0.96 |
0.73 |
|
Hср |
7.57-10.11 |
8.97±0.13 |
0.56 |
7.9-9.5 |
8.78±0.08 |
0.45 |
0.19 |
|
Lc1 |
24.53-30.71 |
28.79±0.30 |
1.29 |
25.8-32.7 |
28.69±0.24 |
1.38 |
0.04 |
|
В % Lc1 |
|||||||
|
Hc |
62.97-81.39 |
70.40±1.22 |
5.33 |
52.2-69.7 |
62.56±0.56 |
3.24 |
0.91 |
|
AO |
26.45-34.23 |
26.96±0.44 |
1.92 |
27.2-35.3 |
30.04±0.31 |
1.83 |
0.29 |
|
O |
16.62-21.77 |
18.88±0.32 |
1.40 |
18.2-23.0 |
20.7±0.18 |
1.07 |
0.74 |
|
Lmx |
36.96-44.33 |
41.99±0.41 |
1.81 |
37.9-44.40 |
42.05±0.23 |
1.34 |
0.02 |
|
Lmd |
47.40-52.52 |
48.89±0.34 |
1.48 |
43.9-52.7 |
48.58±0.38 |
2.21 |
0.36 |
|
iO кожн |
29.83-35.62 |
31.78±0.37 |
1.62 |
27.9-42.5 |
31.04±0.55 |
3.22 |
0.15 |
МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ПАРУСОВИДНОЙ БОРОДАТКИ HISTIODRACO VELIFER (REGAN, 1914) (ARTEDIDRACONIDAE) ИЗ РАЗЛИЧНЫХ МОРЕЙ АНТАРКТИКИ
Парусовидная, или длинноперая бородатка Histiodraco velifer Regan распространена в Южном океане, скорее всего, циркумконтинентально, но пока достоверно известна из морей Уэдделла, Космонавтов, Дейвиса, Содружества и Росса (Eakin, 1990, наши данные). Это – донный вид, населяющий шельф материка и близлежащих островов на глубинах от 30 до 500 м. Он крайне редко встречается в уловах, поэтому в музейных коллекциях мира представлен единичными экземплярами. Литературные сведения о его строении весьма скудны (Regan 1914а, 1914b; Norman, 1937, 1938; Eakin, 1981, 1990; Miller, 1993; Воскобойникова, 1998 и др.). Очень слабо изучена и биология парусовидной бородатки (Eakin, 1990).
Цель настоящей работы – на основе материалов российских и немецких антарктических экспедиций из различных морей Антарктиды (Дейвиса, Космонавтов, Росса, Содружества и Уэдделла) дать более детальное описание вида, включающее признаки внешней морфологии, осевого скелета и сейсмосенсорной системы.
Описание Histiodraco velifer Regan, 1914
Основные счетные признаки: D III–IV, 24–26; А 15–18; Р 17–21; C 20–21; жаберных тычинок на 1-й дуге в наружном ряду 7–12, во внутреннем – 6–10; общее их число на первой дуге 13–22.
Сейсмосенсорная система. В супраорбитальном канале (CSO) обычно 4 поры (5 пор только у 1 экз. ISH 84–1985 слева), в инфраорбитальном (CIO) – 7 пор, в преоперкуломандибулярном – обычно 9 (10 пор у 1 экз. ISH 53–1984 слева), в темпоральном канале (СT) – обычно 6 (5 пор у 1 экз. ISH 53–1984 справа). Корональная коммиссура с 1-ой порой, супратемпоральная комиссура разорвана посередине (нет медиальных сегментов) и содержит обычно по 1-ой поре с каждой стороны головы (по 2 поры у 1-го экз. ЗИН No52135). Преоперкуломандибулярный канал не связан с темпоральным. Изменчивость в числе пор связана с появлением дополнительной сегментной поры (pso1-2) на первом сегменте (=сенсе) супраорбитального канала, появлением 2 дополнительных межсегментных пор (ppm6a и ppm6b) на месте 6-й основной поры преоперкуломандибулярного канала, появлением дополнительной сегментной поры pst0-1 («pso0»=pt3, т.е. это – общая пора между темпоральным каналом и супратемпоральной комиссурой, которую принято относить к CT) и зарастанием последней поры (pt6) темпорального канала. В дорсальной боковой линии (ULL) 15–22 пор, в медиальной (срединной) боковой линии 12–20 «чувствующих» чешуй (т.е. чешуй, несущих крупные свободные невромасты; подробнее см.: Балушкин, 1996).
Рентгенографические данные (n = 8): позвонков до 1-го проксималь-ного птеригиофора ID (=interneurale) – 1–2, до 1-й interneurale IID – 6–7, до 1-го проксимального птеригиофора A (= 1-й interhaemale) – 14. Между спинными плавниками 1–2 interneuralia без лучей; здесь всегда остается 1 межпозвонковый промежуток, в котором нет interneurale. Первый проксимальный птеригиофор анального плавника располагается снизу, на границе туловищного и хвостового отделов позвоночника. Свободных хвостовых позвонков сверху (за последним птеригиофором IID) – 6–7, снизу (за последним птеригиофором A) – 5–7. К уростилярному позвонку причле-няются 4 гипуральные пластинки: parhypurale + нижняя комплексная гипурале (H1+H2) + верхняя комплексная гипурале (H3+H4) + hypurale minimale (H5), или только 3. В последнем случае hypurale minimale частично или полностью сливается с верхней комплексной гипурале, образуя комплекс (H3+H4+Н5). Все гипуральные пластинки слиты с уростилем. В хвостовом плавнике 20–21 луч: 4 верхних краевых + 13 основных + 3–4 нижних краевых. Основные лучи распределяются по гипуральным пластинкам в отношении (сверху вниз) 1–6–4–2; 1–6–5–1 (у рыб с 4-мя пластинками) и 7–4–2 или 7–5–1 (у рыб с 3-мя пластинками). Эпуралий 2, передняя из них по форме сходна с верхним остистым отростком предуростилярного позвонка. Одна пара uroneuralia.
Морфометрия. В процентах SL: антедорсальное расстояние до ID – 34.2–39.6 (36.9); до IID – 40.8–50.2 (46.5); антеанальное расстояние – 62–69.9 (65); высота тела у анального плавника – 14.1–22.5(17.9); длина грудного плавника – 22.5–31.7 (26.9); длина брюшного плавника – 14.1–25 (19.7); длина хвостового стебля – 3.8–9.2 (6.4), его высота – 6.6–8.9 (7.7); длина головы до конца operculum – 37.7–51.8 (42.9); ширина головы у предкрышек – 22.8–30.8 (27.1); высота головы на уровне посттемпоральных гребней – 20.7–26.9 (23.6); в процентах длины головы (Lc1): длина рыла составляет 19.7–30.1 (26.1); диаметр глаза – 24.7–36.2 (30.1); длина верхней челюсти – 31.9–46 (40.6); длина нижней челюсти – 46.6–54.4 (46.9); длина подбородочного усика – 36.2–62.4 (50.3); межглазничное расстояние между костными краями frontalia – 4.8–12.1 (8.7). Данные подсчетов и измерений рыб раздельно для морей Космонавтов, Росса, Моусона, Содружества и Уэдделла приведены в табл. 8.
Подбородочный усик. Подбородочный усик (см. рис. 1) длинный, составляет в среднем около половины длины головы (36.2–62.4% Lc1). Стебель подбородочного усика тонкий, со слабо выраженным терминальным расширением (=эской). Эска составляет от 7.3 до 38.6 % длины усика (в среднем 19.6%), она уже ширины стебля у его основания и лишь немного шире стебля в его дистальной части. Нижней границей эски мы считали то место, где наблюдается заметное сгущение дополнительных придатков (отдельно сидящие удлиненные придатки могут иногда располагаться на стебле значительно ближе к его основанию).
Таблица 8. Счетные и пластические признаки парусовидной бородатки Histiodraco velifer из морей Космонавтов, Росса, Моусона, Содружества и Уэдделла
|
Признаки |
Море Космонавтов |
Море Росса |
Море Моусона |
Море Содружества |
Море Уэдделла |
||||||||||
|
lim |
x±mx |
σ |
lim |
x±mx |
σ |
lim |
x±mx |
σ |
lim |
x±mx |
σ |
lim |
x±mx |
σ |
|
|
D1 |
3-3 |
3±0 |
0 |
3-3 |
3±0 |
0 |
3-3 |
3±0 |
0 |
3-3 |
3±0 |
0 |
3-4 |
3.2±0.2 |
0.4 |
|
D2 |
25-26 |
25.8±0.2 |
0.4 |
24-26 |
25.3±0.4 |
0.8 |
26-26 |
26±0 |
0 |
25-25 |
25±0 |
0 |
25-26 |
25.6±0.2 |
0.5 |
|
A |
17-18 |
17.3±0.2 |
0.5 |
15-17 |
16.5±0.4 |
0.9 |
17-18 |
17.5±0.5 |
0.7 |
16-17 |
16.5±0.5 |
0.7 |
16-18 |
17±0.2 |
0.6 |
|
P |
18-19 |
18.5±0.2 |
0.5 |
17-18 |
17.8±0.2 |
0.4 |
19-19 |
19±0 |
0 |
17-18 |
17.5±0.5 |
0.7 |
18-21 |
19.1±0.4 |
1 |
|
Sp.br. ant. |
7-12 |
9.1±0.5 |
1.4 |
8-11 |
9±0.6 |
1.2 |
8-10 |
9±1 |
1.4 |
9-9 |
9±0 |
0 |
7-11 |
8.7±0.6 |
1.5 |
|
Sp.br. post. |
6-10 |
7.6±0.5 |
1.3 |
6-7 |
6.8±0.2 |
0.4 |
6-8 |
7±1 |
1.4 |
7-8 |
7.5±0.5 |
0.7 |
6-9 |
6.9±0.4 |
1.1 |
|
В % стандартной длины (SL) |
|||||||||||||||
|
Lc1 |
39.5-43.4 |
41.5±0.4 |
1.3 |
40.5-44.1 |
42±0.7 |
1.4 |
37.7-51.8 |
44.8±7 |
10 |
43.1-49.4 |
46.3±3.2 |
4.4 |
41-46.8 |
43.7±0.7 |
1.9 |
|
Hc |
22.3-25.8 |
24.3±0.4 |
1.1 |
20.7-25.5 |
23.4±1 |
2 |
22.5-22.6 |
22.6±0 |
0.1 |
25.7-26.9 |
26.3±0.6 |
0.9 |
21.1-25.4 |
22.4±0.6 |
1.6 |
|
Wc |
24.8-30.7 |
28.4±0.7 |
1.9 |
24.1-30.3 |
27±1.3 |
2.5 |
25.3-29.1 |
27.2±1.9 |
2.7 |
29.6-30.8 |
30.2±0.6 |
0.8 |
22.8-26.5 |
24.5±0.5 |
1.3 |
|
H |
17.9-22.5 |
19.7±0.5 |
1.3 |
16.4-18 |
17±0.3 |
0.7 |
17.5-17.9 |
17.7±0.2 |
0.3 |
17.8-20.4 |
19.1±1.3 |
1.8 |
14.1-18.1 |
15.9±0.6 |
1.4 |
|
aD1 |
34.2-38.2 |
36.1±0.4 |
1.3 |
36.3-39.5 |
37.2±0.7 |
1.4 |
34.2-37.7 |
35.9±1.7 |
2.5 |
37.1-38.9 |
38±0.9 |
1.3 |
36.5-39.7 |
37.9±0.4 |
1.1 |
|
aD2 |
40.8-47.7 |
45.4±0.7 |
1.9 |
46.5-50.3 |
48±0.7 |
1.5 |
43.9-45.3 |
44.6±0.7 |
1 |
46.5-48.5 |
47.5±1 |
1.4 |
44.6-49.6 |
47.4±0.8 |
2.1 |
|
aA1 |
62-66.6 |
64.1±0.5 |
1.6 |
62.6-66.4 |
64.7±0.8 |
1.7 |
62.2-69.9 |
66.1±3.9 |
5.5 |
62.8-67.3 |
65.1±2.3 |
3.2 |
63.7-69 |
66.2±0.7 |
1.8 |
|
LD1 |
20.7-30.8 |
24.3±1.2 |
3.5 |
17.5-25.4 |
21.7±1.4 |
2.8 |
15.3-22.4 |
18.9±3.5 |
5 |
22.7-24.5 |
23.6±0.9 |
1.3 |
21.8-34 |
26.3±1.8 |
4.8 |
|
LD2 |
26.4-42.7 |
35.3±1.9 |
5.8 |
25-31.8 |
29.5±1.3 |
2.7 |
21.5-37 |
29.3±7.8 |
11 |
30-30.6 |
30.3±0.3 |
0.5 |
24.1-43.3 |
32.1±2.6 |
7 |
|
LP |
23.8-31.7 |
28.5±0.8 |
2.4 |
23.9-25.8 |
25±0.4 |
0.8 |
23.4-24.4 |
23.9±0.5 |
0.7 |
24.1-25.7 |
24.9±0.8 |
1.1 |
22.5-29.3 |
27.6±0.9 |
2.3 |
|
LV |
16.8-22.5 |
19.2±0.6 |
1.6 |
14.2-17.8 |
16.8±0.8 |
1.5 |
14.3-20.9 |
17.6±3.3 |
4.7 |
17.5-19.7 |
18.6±1.1 |
1.5 |
21.4-25 |
23±0.5 |
1.2 |
|
Lcp |
5.4-8.5 |
6.8±0.3 |
1 |
3.8-9.3 |
6.6±1 |
2 |
5.1-5.7 |
5.4±0.3 |
0.5 |
5.7-7.3 |
6.5±0.8 |
1.1 |
4.9-6.9 |
5.9±0.3 |
0.7 |
|
Hcp |
7-8.9 |
7.8±0.2 |
0.6 |
7.2-8.4 |
7.7±0.2 |
0.5 |
6.7-8.6 |
7.6±1 |
1.4 |
7.3-8.7 |
8±0.7 |
1 |
6.9-7.9 |
7.4±0.1 |
0.3 |
|
aO |
19.7-30.1 |
26.2±1.1 |
3.2 |
26.7-29.2 |
28.2±0.5 |
1 |
24-26.2 |
25.1±1.1 |
1.5 |
25.6-29.1 |
27.4±1.8 |
2.5 |
23.2-26.7 |
24.7±0.5 |
1.4 |
|
O |
28.1-36.2 |
31.2±0.8 |
2.4 |
27.2-30.8 |
29.8±0.8 |
1.5 |
24.7-26.4 |
25.5±0.9 |
1.2 |
31-31.2 |
31.1±0.1 |
0.1 |
27.8-32.4 |
29.8±0.6 |
1.5 |
|
Lbarb |
36.2-60.6 |
52.6±2.2 |
6.5 |
45.6-58.1 |
52.9±2.3 |
4.6 |
42.4-62.4 |
52.4±10 |
14.2 |
44.1-54.7 |
49.4±5.3 |
7.5 |
42.2-51.4 |
45.7±1.1 |
3 |
|
iO |
4.8-12.1 |
9.2±0.8 |
2.5 |
7.5-9.1 |
8.3±0.4 |
0.7 |
7.9-9.2 |
8.6±0.7 |
1 |
8.3-9.3 |
8.8±0.5 |
0.7 |
6.8-9.8 |
8.1±0.6 |
1.5 |
|
Lmx |
35.4-46 |
40.6±0.9 |
2.8 |
40-43.8 |
41.4±0.7 |
1.4 |
31.9-38.6 |
35.2±3.4 |
4.8 |
37.2-40.5 |
38.8±1.7 |
2.3 |
41.4-43.8 |
42.2±0.3 |
0.9 |
|
Lmd |
42.1-54.4 |
47.3±1.2 |
3.6 |
45.3-47.9 |
46.5±0.5 |
1 |
36.6-45.3 |
41±4.4 |
6.2 |
42.4-43.6 |
43±0.6 |
0.9 |
46.3-53.5 |
49.6±1 |
2.6 |
|
Md-Mx |
2.2-5.5 |
4±0.3 |
1 |
0.5-3.9 |
2.1±0.7 |
1.5 |
1.9-8.4 |
5.2±3.3 |
4.6 |
1.7-3 |
2.4±0.6 |
0.9 |
5.1-7.6 |
6.2±0.4 |
0.9 |
|
|
|
|
|
|
|
В % длины усика (Lbarb) |
|
|
|
|
|
|
|||
|
Le |
7.3-31.7 |
18.5±2.3 |
6.9 |
18.5-38.6 |
24.9±4 |
8.1 |
10.8-13 |
11.9±1.1 |
1.6 |
15.7-29.2 |
22.5±6.8 |
10 |
- |
- |
- |
Длина и форма придатков у рыб значительно варьирует: они могут быть в виде простых коротких усиков обычной (как это изображено у Андрияшева, 1967, рис. 1, 3) или серповидной (клешнеобразной) формы, или более длинных придатков, несущих мелкие вторичные отростки. У одного из экземпляров из моря Росса (рис. 4, а, 1) стебель усика раздвоен, и каждая его ветвь несет длинные отростки, снабженные вторичными папиллами, длина которых уменьшается дистально. Стебель усика обычно густо покрыт мелкими папиллами. Усик слабо пигментирован, у части экземпляров хорошо сохраняется темное пигментное пятно в дистальной части эски. Особенностей строения усика, указывающих на наличие полового диморфизма, выявлено не было (рис. 4). Ранее у близкого вида Dolloidraco longedorsalis также не было найдено связи между строением усика и полом (Eastman, Eakin, 2001). Функция подбородочного усика у рыб сем. Artedidraconidae пока окончательно не выяснена. Согласно одним данным (MacDonald, Montgomery, 1991; Janssen et al., 1993), усик используется как привлекающая приманка и как соматосенсорный орган, помогающий обнаруживать добычу. Японским ученым (Iwami et al., 1996), проводившим наблюдения над рыбами, помещенными в аквариум, не удалось подтвердить факт использования усика в качестве приманки. Они считают, что он выполняет лишь тактильную функцию, позволяющую определять присутствие и положение добычи. Гистологические исследования показали, что усик антарктических бородаток лишен вкусовых рецепторов (вкусовых почек) (Eastman, Eakin, 2001).
Окраска (в спирте). Голова более светлая сверху и снизу (включая жаберные перепонки). Щеки, жаберные крышки и брюхо темно-коричневые. Темные коричневые пятна неправильной формы на слезных костях и челюстях. Дистальные концы верхней челюсти светлые. На боках тела в верхней части обычно 4 широкие темные поперечные полосы, частично заходящие на спинные плавники: 1-я полоса находится под первым спинным плавником, 2-я – под первыми 5–6 лучами IID, 3-я – под 9–11 лучами IID и наиболее широкая 4-я полоса – под последними лучами IID (она может распадаться на 2-3 частично изолированные друг от друга полоски). В нижней части туловища поперечные полосы разбиваются на отдельные пятна, образующие сложный (иногда – близкий к шахматному) узор. Спинной и анальный плавники с темными пятнами, образующими косые полосы. Грудной и брюшной плавники с темными пятнами, формирующимися в неправильной формы поперечные полосы. Крупные темные пятна на основании грудного плавника и хвостовом стебле (включая основания лучей хвостового плавника). Хвостовой плавник с темной широкой поперечной полосой посередине, концы лучей светлые.

Рис. 4. Строение подбородочных усиков у экземпляров Histiodraco velifer Regan из различных морей Южного океана. Обозначения (номер на рисунке – коллекционный номер экземпляра, в скобках – порядковый номер экземпляра в серии, стандартная длина тела): а) - самки: 1 – No52134(3), 119 мм; 2 – No52135(9), 131мм; 3 – No52135(6), 142 мм; 4 – No52134(2), 133 мм; 5 – No46459, 159мм; 6 – No52132(2), 157мм; 7 – No52135(1), 152 мм; 8 – No52135(7), 136,5 мм; 9 – No52134(1), 157 мм; b) - самцы: 1 – No52135(4), 137 мм; 2 – No52132(1), 117 мм; 3 – No52135(5), 147 мм; 4 – No52135(2), 138 мм; 5 – No52135(8), 146 мм; 6 – No52133, 158 мм SL; c) - ювенильные особи: 1 – No52135(3), 117 мм; 2 – No37500, 123 мм.
Внутривидовая изменчивость. Несмотря на то, что нами исследована самая представительная коллекция Histiodraco velifer, малые размеры выборок из различных морей Южного океана оставляют пока открытым вопрос о наличии морфометрической обособленности их друг от друга. Кластерный анализ общего сходства, проведенный по 25 признакам (четыре счетных и 21 пластический), показывает отсутствие строгого группирования рыб по их географической принадлежности (рис. 5). Не выявлено нами также сколько-либо заметных различий рыб из морей Космонавтов, Росса, Моусона, Содружества и Уэдделла в качественных признаках (окраска, строение позвоночника, сейсмосенсорной системы, подбородочного усика и др.). Воскобойникова (1998) обратила внимание на расхождение в числе жаберных тычинок у описываемого ею малька из моря Уэдделла с данными Икина (Eakin, 1990) (r.br. 1+7 vs 1-3 + 14-17), объясняя это неполнотой развития тычинок у малька. В действительности никакого расхождения данных у этих авторов нет, поскольку указанное Р. Икиным общее число тычинок на первой дуге (sp.br.ant. + sp.br.post.) Воскобойникова ошибочно приняла за число тычинок только в наружном (внешнем) ряду (sp.br.ant.) первой дуги. Судя по ранее опубликованным данным Икина (Eakin, 1981: 88), число тычинок в наружном ряду 1-й дуги у исследованных им рыб из моря Росса составляло 1 + 0-1 + 7-9, что хорошо соответствует таковым у малька из моря Уэдделла.
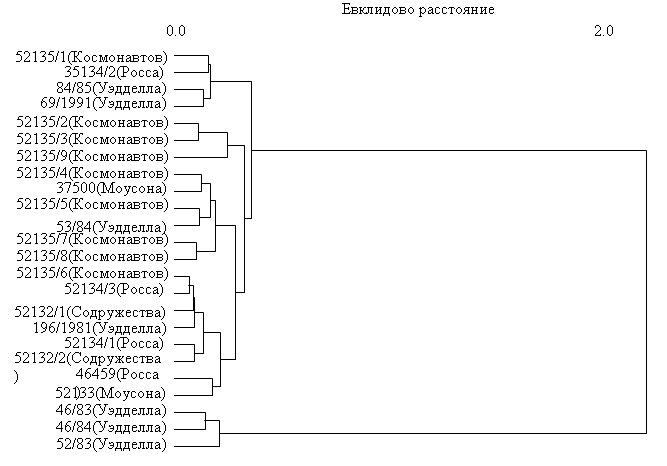
Рис. 5. Дендрограмма различий экземпляров Histiodraco velifer по 25 морфологическим признакам из морей Космонавтов, Моусона, Росса, Содружества и Уэдделла. Алгоритм кластеризации: Ward Minimum Variance Method.
ЛИТЕРАТУРА
Андрияшев А.П. 1967. Обзор рыб-бородаток рода Pogonophryne Regan (Pisces, Harpagiferidae) с описанием пяти новых видов из Восточной Антарктики и Южных Оркнейских островов // Исслед. фауны морей. Т. 4 (12). Л.: Наука. С. 389–412. – 1986. Общий обзор фауны донных рыб Антарктики // Морфология и распространение рыб Южного океана / Тр. Зоол. ин-та АН СССР. Т. 153. С. 9–45.
Андрияшев А.П., Токарев А.К. 1958. Ихтиофауна // Тр. компл. Антарк. эксп. АН СССР, описание экспедиции д/э «Обь» 1955–1956. М. С. 195–207.
Балушкин А.В. 1976. Обзор группы видов "larseni" рода Notothenia Rich. // Вопросы ихтиологии. Т. 16. Вып. 1. С. 3–15. – 1984. Морфологические основы систематики и филогении нототениевых рыб. Л.: Изд-во Зоол. ин-та АН СССР, 141 с. – 1986. Морфометрическая характеристика зеленой нототении Gobionotothen gibberifrons (Lonnberg, 1905) из различных районов Западной Антарктики // Тр. Зоол. ин-та АН СССР. Т. 153. С. 72–79. – 1991. Обзор зеленых нототений рода Gobionotothen Balushkin (сем. Nototheniidae) Антарктики и Субантарктики // Вопр. ихтиологии. Т. 31. Вып. 4. С. 531–542. – 1992. Классификация, родственные связи и происхождение семейств нототениоидных рыб подотряда Notothenioidei // Вопр. ихтиологии. Т. 32. Вып. 2. C. 3–19. – 1996. Строение и эволюция туловищных боковых линий нототениоидных рыб (Notothenioidei, Perciformes) // Вопр. ихтиологии. Т. 36. No 4. С. 437–447. – 2002. Повторное видообразование у рыб отдаленных островов Южного океана: Тез. докл. научной конференции "Исследования и охрана окружающей среды Антарктики" (Санкт-Петербург, 13–15 ноября 2002 г.) / Экспресс-информация Гос. научн. центра Российской Федерации – Арктич. и Антарктич. научно-исслед. ин-та. СПб. Вып. 15. С. 16–17.
Балушкин А.В., Федоров В.В. 2002. Современные исследования фауны рыб Южного океана // Известия Зоологического института РАН. No 4. 32 с.
Берг Л.С. 1977. Труды по теории эволюции. Л.: Наука, 387 с.
Воскобойникова О.С. 1998. Молодь Histiodraco velifer (Artedidraconidae, Notothenioidei) из моря Уэдделла (Антарктика) // Вопр. ихтиологии. Т. 38. No 2. С. 273–274.
Воскобойникова О.С., Балушкин А.В. 1987. О таксономическом статусе Gobionotothen angustifrons sandwichensis (Nybelin, 1947) (Nototheniidae) // Тр. Зоол. ин-та АН СССР. Т. 162. С. 100–107.
Кузнецов Б.А. 1936. О некоторых закономерностях распространения млекопитающих по европейской части СССР // Зоол. журн. Т. 15. Вып. 1. С. 96–127.
Природина В.П. 2000. О систематическом положении литоральных и глубинных видов рода Harpagifer (Harpagiferidae, Notothenioidei) от острова Маккуори с описанием двух новых видов // Вопр. ихтиологии. Т. 40. No 4. С. 438–445. – 2002. Переописание литоральных и глубинных видов рода Harpagifer (Harpagiferidae, Notothenioidei) из индоокеанского сектора Южного океана с описанием нового вида // Вопр. ихтиологии. Т. 42. No 6. С. 731–743.
Смирнов И.С. 1990. Фауна змеехвосток (Echinodermata: Ophiuroides) Южного океана и влияние течения Западных Ветров на распространение офиур // Океанологические исследования Южного океана. Междуведомственный сборник научных трудов. Л.: Изд-во ЛГУ. С. 165–188. – 1992. Типы ареалов змеехвосток (Echinodermata: Ophiuroides) Южного океана // Физическая океанология и проблемы биологической продуктивности. Междуведомственный сборник. СПб: Изд-во СПбГУ. С. 136–165.
Танкевич П.Б. 1986. Морфометрическая характеристика кергеленской мраморной нототении Notothenia rossii rossii Richardson из Индийского сектора Южного океана // Вопр. ихтиологии. Т. 26. Вып. 2. С. 328–332.
Шуст К.В. 1998. Рыбы и рыбные ресурсы Антарктики. М.: ВНИРО, 161 с.
Якубовски М. 1971. Морфологические особенности системы органов боковой линии у представителей рода Notothenia Rich. и других родов семейства Nototheniidae (Pisces) // Вопр. ихтиологии. Т. 11. Вып. 4. С. 595–601.
Andersen N.C. 1984. Genera and subfamilies of the family Nototheniidae (Pisces, Perciformes) // Steenstrupia. Vol. 10. No 1. P. 1–34.
Andersen N.C., Hureau J.-C. 1979. Proposition pour une nouvelle classification des Nototheniidae (Pisces, Perciformes, Nototheniidae) // Cybium. Ser. 3. No 6. P. 47–53.
Balushkin A.V. 1989. Morphological bases of the systematics and phylogeny of the nototheniid fishes. Oxonian Press Pvt. Ltd., New Delhi-Calcutta. 153 p. – 2000. Morphology, classification, and evolution of notothenioid fishes of the Southern Ocean (Notothenioidei, Perciformes) // J. Ichthyology. Vol. 40. Suppl. 1. P. S74–S109.
Balushkin A.V., Fedorov V.V. 2001. Check list of Antarctic fishes // VII SCAR International Biology Symposium “Antarctic Biology in a Global Context”. Sec. 5. P. 1.
DeWitt H.H., Heemstra P.S., Gon O. 1990. Nototheniidae. P. 279–331 // Fishes of the Southern Ocean (Gon O., Heemstra P.C, eds). J.L. B. Smith Inst. Ichthyol. Grahamstown, 461 p.
Eakin R.R. 1981. Osteology and relationships of the fishes of the Antarctic family Harpagiferidae (Pisces, Notothenioidei) // Biology of the Antarctic Seas IX (Kornicker L.S., ed.) / Antarc. Res. Ser. Vol. 31. P. 81–147. – 1990. Artedidraconidae. P. 332–356 // Fishes of the Southern Ocean (Gon O., Heemstra P.C., eds.). J.L.B. Smith Inst. Ichthyol. Grahamstown, 461 p.
Eastman J.T., Eakin R.R. 2001. Mental barbell and meristic variation in the Antarctic notothenioid fish Dolloidraco longedorsalis (Perciformes: Artedidraconidae) from the Ross Sea // Polar Biol. No 24. P. 729–734.
Gon O., Klages N.T. 1988. The marine fish fauna of the Sub-Antarctic Prince Edward Islands // S. Afr. J. Antarct. Res. Vol. 18. No 2. P. 32–54.
Hardy G. S. 1988. A revision of Bovichtus Cuvier, 1831 (Pisces: Bovichthyidae) from Australasia, with description of a new deepwater species from the New Zealand Subantarctic // J. Nat. Hist. Vol. 22. P. 1639–1655.
Hureau J.-C. 1970. Biologie comparee de quelques Poissons antarctiques (Nototheniidae) // Bull. Inst. Oceanogr. Monaco. Vol. 68. No 1391. P. 1–244.
Iwami T., Numanami H., Naito Y. 1996. Behavior of three species of the family Artedidraconidae (Pisces, Notothenioidei), with reference to feeding // Proc. NIPR Symp. Polar Biol. No 9. P. 225–230.
Janssen J., Jones W., Slattery M. 1993. Locomotion and feeding responses to mechanical stimuli in Histiodraco velifer (Artedidraconidae) // Copeia. No 3. P. 885–889.
MacDonald J.A., Montgomery J.C. 1991. The sensory biology of notothenioid fish // Biology of Antarctic fish (di Prisco G., Maresca B., Tota B., eds.). Berlin: Springer. P. 145–162.
Miller R.G. 1993. A History and Atlas of the fishes of the Antarctic Ocean. Foresta Institute for Ocean and Mountain Studies. Carson City, Nevada, 792 p.
Norman J.R. 1937. Fishes // Report B.A.N.Z. Antarctic Res. Exp. 1929–1931. Ser. B. Vol. 1. Part 2. P. 51–88. – 1938. Coast fishes. Part 3. The Antarctic Zone // Disc. Rep. Vol. 18. P. 1–105.
Nybelin O. 1947. Antarctic fishes // Sci. Res. Norweg. Antarct. Exp. 1927–1928. Oslo. Vol. 2. No 26. P. 1–76.
Regan C.T. 1914a. – Diagnoses of new marine fishes collected by the British Antarctic ("Terra Nova") Expedition // Ann. Mag. Nat. Hist. Ser. 8. Vol. 13. No 73. P. 11–17. – 1914b. Fishes // Nat. Hist. Rep. Brit. Antarct. ("Terra Nova") Exp. 1910. Zool. Vol. 1. No 1. P. 1–54.
Richardson J. 1844. Ichthyology of the voyage of the H.M.S. Erebus & Terror, under the command of Captain Sir James Clark Ross, R.N., F.R.S. London, 1844–1848. Vol. 2. P. I-VII + 1–139.
Sosinski J., Skora K.E. 1988. Results of investigation R/V Profesor Siedlecki of the state of fish stocks in the Scotia Arc area in the 1986/1987 seasson // Biul. Morsk. Inst. Ryb. No 3–4 (107–108). P. 31–43.
Worner F.G., James R. 1981. Early life history stages of the Antarctic fish Notothenia gibberifrons Lonnberg, 1905 // Papp. P.-V. Reun. Cons. Int. Explor. Mer. No 178. P. 196.