
|

|




|
|
|
|
|
|
|
|
The proposed scheme of taxonomic composition of the order Coleoptera is based on the scheme published in 1991 (Kirejtshuk, 1991) with changes made in accordance with new materials and interpretations which be discussed in some future.

|
Fig. 1. Taxonomic composition and distribution of basic coleopterous groups in the paleontologic chronicle. |
The order Coleoptera is rather unique in relation to the great number and alignment extinct and recent families. At present 166 families are recognized for the recent fauna. 67 families are known from the Palaeozoic and Mesozoic eras and only 19 of them became extinct before the Caenozoic era (including all palaeozoic families) (Ponomarenko, 1995). Remnants of recent families of other orders usually go down not lower than the Palaeogene, however coleopterous families reach the Cretaceous, sometimes Jurassic or even Triassic deposits. In other words Coleoptera presents the group, historical development of which was going on less dramatically in contrast to those of other orders of the class, unless than other orders of terrestrial animals. Somewhat similar situation among insects can be traced only in the order Heteroptera, where 40 from about 50 recent families are recorded from deposits (9 from the Mesozoic deposits) and there were described only 16 extinct families (Principles of palaeontology, 1962 - Rodendorf, ed.).
Almost all coleopterous superfamilies were formed before or during the Cretaceous crises. Stability of many coleopterous groups in time can be explained by their origin as inhabitants in subcortical and similar sites of the xeromorphous flora of Gymnosperms at the Rubicon of Carboniferous and Permian (Ponomarenko, 1969). Basic phyletic transformations were at these sites (suborders Archostemata and Polyphaga, infraorder Cucujiformia) or defined by shifts of part of ancient beetles to other localities (suborders of Adephaga and Myxophaga - in continental basins; infraorders Staphytliniformia and Elateriformia - to wood after long period of decay; infraorder Curculioniformia - to generative organs of Gymnosperms - Kirejtshuk, 1992). A record of longivity without great transformation was set up by representatives of the suborder Archostemata (Kirejtshuk, 1999), because this suborder seemed to be formed in connection with adaptation of their larvae to boring of wood and maintaining a type of ontogenesis initial for the first beetles (representatives of Archostemata appeared in the Triassic and remnants of insects boring in petrified trunks of Gymnosperms are known from this period - Crowson, 1981).
Scales and rates of phyletic changes in a certain sense are in direct proportion to intensivity and duration of influence of changing environment (Starobogatov, Levchenko, 1993). Active movements of larvae in more or less open localities promote maintainance of a steady tendency to embryonization of development before emergence of larva from egg and imaginigation of larval differentiation (suborder Adephaga and infraorder Staphyliniformia), while inactive mode of life in substrate which present a food brings to a progresive desembryonyzation of development in egg and shifting of differentiation in pupa (infarorder Curculioniformia and superfamily Chrysomeloidea) (Tikhomirova, 1991; Kirejtshuk, 1994, 1996). The groups with a considerable longivity of both larval and imaginal life are frequently characterized by gerontomorphosis and predominance of anaboly in transformations of imaginal differentiation, while the forms with short larval development and not long imaginal life usually show paedomorphoses in structure of different organs and it is reasonable to admit abbreviations as a more probable mode of their evolution (Ponomarenko, 1983; Kirejtshuk, 1994, 1996). A constant intensification of most processes in the biosphere in time explains prevalence of the groups with traced paedomorphoses among beetles. Shortening of definitive differentiation conducts to losses of traces of latest phyletic connections and difficulties in reconstruction of phylogenetic links of many groups.
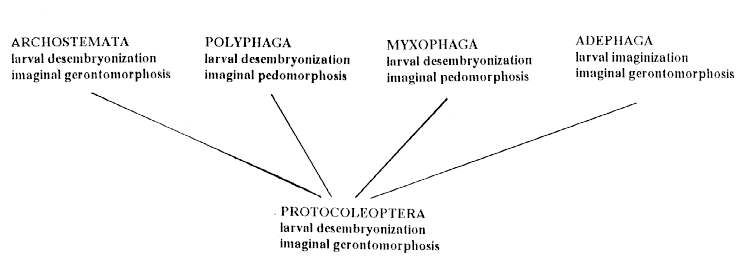 |
Fig. 2. Types of ontogenesis in different suborders of Coleoptera. |
Palaeontological chronicle increasingly more concentrated by fossil records gives a good base for phylogenetic interpretations. However, a tradition to consider a phylogeny as a divergent differentiation from a generalized ancestor usually remains by ideological base for these interpretations. Sequence of divergent events, produced from analysis of structural characters would be admissible only at absence of paedomorphoses with losses of the definitive differentiation. Due to paedomorphous losses each analysis throws the analized groups to generalized ancestors in proportions to the deepth and scope of paedomorphic transformations. It can be shown in the cladograms elaborated for the infraorder Elateriformia (Lawrence, 1988; Lawrence et al., 1995). Data from other fields of knowledge should be used to correct the results of character analysis. The most important of them are ontogenetics and palaeontological chronicle. The ontogenetic method gives as good results in such corrections as it did before, if to take into consideration a plasticity of mechanisms of ontogenetic transformations in a normal regularity of individual development (Tikhomirova, 1991). "Method of accumulation of appearances" has its limits of solutions for different groups, including some restrictions in interpretations of results (Dmitriev, Zherihin, 1988), although it allows to make probable supposions on dinamics of appearance of groups and to correct the data after analysis of structural features. The biochemical method received wide acceptance in the last decades seems to promise a lot of new results, however it is probable that the applicability of it in phylogenetic studies will be more restricted in comparison with the perspectives seeming at the present. Establishment of sequences in structure of DNA will give a very valuable materials for the analyses, which are now used for analyses of structural traits, but they will scarcely change solvable possibilities of phylogenetics in principle.
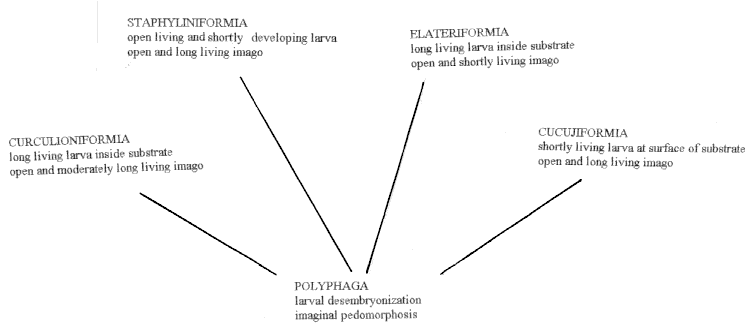 |
Fig. 3. Mode of life in different groups of the suborder Polyphaga. |
Mode of life can be regarded as one of integrated inheritable features, connected with inherited predisposition and conservatism of ontogenegis. Modifications of it look like more significant for a formation of a taxonomic syndrome of a group of related species than changes in structural characters in accordance with particular adaptations (Linnean principle, according to which a group defines the characters). Thus, study of evolution of mode of life can give a valuable material for corrections of phylogenetic hypotheses and for reconstruction of evolution of a group (Kirejtshuk, 1994, 1996).
A.G. Kirejtshuk, June 2000
Crowson R. A.
The biology of Coleoptera. - London-N.Y.-
Toronto-Syney-San Francisco: Academic Press - 1981 - 802 pp.
Dmitriev V.Yu., Zherihin V.V.
Izmenenie raznoobraziya
semeystv po dannim metoda nakopleniya poyavleniy // V kn.:
pod red. A.G. Ponomarenko. Melovoy biotsenoticheskiy
krizis i evolyutsiya nasekomykh. - M.: Nauka - 1988 - S. 208-
215.
Kirejtshuk A.G. 1992
Evolution of mode of life as the basis for division of the beetles into
groups of high taxonomic rank. In: Advances in Coleopterology (Eds.
M.Zunino, X.Belles, M.Blas). Barcelona. P. 249-262.
Kirejtshuk A.G. 1994
Sistema, evolyutsiya obraza zhizni i
filogeniya otryada zhestkokrylykh (Coleoptera). I. // Entom.
obozr. - 1994 - T. 73, Vip. 2 - S. 266-300.
Kirejtshuk A.G. 1996
Sistema, evolyutsiya obraza zhizni i
filogeniya otryada zhestkokrylykh (Coleoptera). II. // Entom.
obozr. - 1996 - T. 75, Vip. 1- S. 39-62.
Kirejtshuk A.G. 1999
Sikhotealinia zhiltzovae (Lafer, 1992) - recent
representative of the Jurassic coleopterous fauna (Coleoptera,
Archostemeta, Jurodidae) // Proc. Zool. Inst. RAS - 1999 - Vol.
281 - 21-26.
Lawrence J.F. 1988
Rhinorhipidae, a new beetle family from
Australia, with comments on the phylogeny of the Elateriformia
// Invertebr. Taxonomy - 1988 - Vol. 2 (1987) - P. 1-53.
Lawrence J.F., Nikitsky N.B.& Kirejtshuk A.G. 1995
Phylogenetic
position of Decliniidae (Coleoptera: Scirtoidea) and comments
on the classification of Elateriformia (sensu lato) // in: J.
Pakaluk & S.A. S'lipin'ski (eds.). Biology, Phylogeny, and
Classification of Coleoptera. - Papers celebrating the 80th
Birthday of Roy A. Crowson. - Warszawa: Muz. Inst. Zool.
PAN - 1995 - Vol. 1 - P. 375-411.
Osnovi paleontologii. 1962
Spravochnik dlya paleontologov i
geologov SSSR. Chlenistonogie: Trakhejnye i khelitserovye,
pod red. Rodendorfa B.B. - M.: Izd. AN SSSR - 1962 - 560
ss.
Ponomarenko A.G. 1969
Istoricheskoe razvitie zhestkokrylykh-arhostemat.
- Tr. Paleont. in-ta AN SSSR - 1969 - T. 125 -
233 ss.
Ponomarenko A.G. 1983
Istoricheskoe razvitie zhestkokrylykh
nasekomykh. Avtoref. diss. st. dokt. biol. nauk - M. - 1983 -
47 ss.
Ponomarenko A.G. 1995
The geological history of beetles // in: J.
Pakaluk & S.A. S'lipin'ski (eds.). Biology, Phylogeny, and
Classification of Coleoptera. Papers celebrating the 80th
Birthday of Roy A. Crowson. - Warszawa: Muz. Inst. Zool.
PAN - 1995 - Vol. 1 - P. 155-172.
Starobogatov Ya.I., Levchenko V.F. 1993
Ekotsentricheskaya
kontseptsiya makroevolyutsii // ZH. obsh'. biol. - 1993 - Tom 54,
vip. 4 - S. 389-407.
Tikhomirova A.L. 1991
Perestroyka ontogeneza kak mehanizm
evolyutsii nasekomykh. - M.: Nauka - 1991 - 168 pp.