
|

|



|
|
|
|
|
|
|
|
Программа общей систематики.
Настоящая статья представляет собой развернутую программу по предмету, который еще не оформился в самостоятельную дисциплину, но потребность в чем, безусловно, чувствуется, и ряд работников как у нас (назову E.С. Смирнова, П.В. Трентьева, С.Я. Парамонова, В.H. Беклемишева), так и за границей пробуют свои силы или по разработке отдельных частей этой дисциплины, или по сводке того, что имеется в этом направлении. Как правильно указывают многие систематики, у нас в сущности нет систематики как отдельной дисциплины. То, что называется систематикой, есть изложение системы животных или растений, а не общее учение о системе, каковым должна быть общая систематика.
Мое предположение написать книгу об общей систематике должно служить завершением тех мыслей, которые впервые появились у меня, когда я в возрасте 13-14 лет впервые начал определять насекомых и когда я натолкнулся на частый источник ошибок, заключающийся в том, что при ошибке в какой-нибудь тезе мы попадаем в обоих разветвлениях дихотомически таблиц на совершенно одинаковые противоположения. Это общеизвестное положение, нашедшее свое выражение и в законе аналогичной изменчивости Уолша (В.D. Walsh), и в закон гомологических рядов H.И. Вавилова, ясно показывает инадэкватность дихотомических таблиц, и к этому придется вернуться позднее.
Первой попыткой наметить программу работы в отношении реконструкции системы с моей стороны была статья "О форме естественной системы организмов" (1923), естественным продолжением и расширенным изданием которой является настоящая статья.
Необходимость реформы систематики и построения особой дисциплины - общей систематики-вытекает как из расширенного понимания задач общей систематики, так и из ряда практических трудностей систематики. В отношении расширенного понимания могу указать, что вместо двух задач - (1) помощи в определении видов и (2) разъяснения смысла систематических сходств и различий, решаемых обычно в смысле отождествления систематики с филогенией, - мы выдвигаем еще третью задачу: построение рациональной системы, т. е. такой, форма и структура которой вытекала бы из некоторых общих принципов, как это делается в системе математических кривых, форм симметрии в кристаллографии, периодической системе в химии, системе органических соединений и т. д. Сообразно с этим вторая задача не может считаться решенной в смысле отождествления систематики с филогенией, и философское понимание системы может приобрести совсем новый смысл. Вторая и третья задачи составляют область общей теоретической систематики, практические же вопросы по построению наиболее удобных таблиц и других средств для решения первой задачи составляют область общей практической систематики. В этой программе я буду касаться преимущественно задач практической систематики как наиболее актуальных. Кроме того, планомерное решение задач теоретической систематики совершенно невозможно без значительного усовершенствования практической систематики и без получения первых существенных результатов коренной реформы этой дисциплины.
Необходимость реформы прежде всего практической систематики вытекает из следующих основных соображений и групп фактов.
ОСНОВАНИЯ ДЕФЕКТОВ СОВРЕМЕННОЙ СИСТЕМАТИКИ
Об этих основаниях уже отчасти говорилось в моей статье о форме естественной системы организмов. Их можно разбить на три основные группы: логические, натурфилософские и мeтодические.
В отношении общего понимания системы продолжает господствовать формально логический подход: всякая система мыслится как иерархия, причем каждый вид (в логическом смысле) определяется ближайшим родом и специфическим отличием (per genus proximus et per differentiam specificam). Человек, незнакомый с фактической биологической систематикой, несказанно удивлен, когда при описании видов, родов и т.д. не находит четких различий, а данные различия всегда обычно сопровождаются оговоркой, и чем естественнее группа, тем труднее ее определить. На это уже давно обратил внимание Е.С. Смирнов4 правильно указавший, что истинным принципом при группировке видов естественных единиц является не формально логический принцип подчинения роду и специфических отличий, а объединение по конгрегационному принципу, т. е. естественной группой называется такая группа, которая состоит из элементов наиболее сходственных, хотя бы крайние члены такой группы и не включали бы в себя ни одного общего признака, т. е. если даже такую группу и невозможно характеризовать никаким общим признаком (ракообразные, насекомые и т. д.). Например, виды группы могут характеризоваться такими признаками: 1) а, б, в, г, д; 2) б, в, г, д, е; 3) г, д, е, ж; 4) д, е, ж, з; 5) е, ж, з, и; 6) ж, з, и, к. Шестой вид не имеет ни одного общего признака с первым, а потому их никак нельзя включить в один род по формально логической связи, но по конгрегационному способу вся группа видов образует естественный род. Это же, между прочим, приложимо к такому понятию, как гомология в отношении конечностей, образующихся из определенных миотомов. Известно, что в сравнительно-анатомическом ряду зачаток конечности может мигрировать и могут быть случаи двух гомологических органов, не имеющих ни одной гомологической части. Конгрегационный принцип касается структуры системы, оставляя в стороне вопрос об ее форме; как было уже указано в моей статье, мы имеем право различать по крайней мере три основные формы системы: а) иерархическую, б) комбинативную, в) коррелятивную. Примером комбинативной системы может быть многообразная комбинация различных независимых генов при наследовании по Менделю, примером коррелятивной - периодическая система элементов. Размышляя о различных системах (математических кривых, форм кристаллов, органических соединений, организмов, болезней и т. д.), можно, пожалуй, прийти к выводу, что настоящей иерархической системы в природе вообще не существует. В этом смысле мнение (если не ошибаюсь, Гете; кажется, об этом есть у Радля5 в его истории биологических учений) о том, что всякая система искусственна, заключает известную долю истины. Гете под системой понимал, как большинство людей, иерархическую систему и не представлял себе, что мыслима система, построенная не на иерархическом принципе. А так как он, вероятно, наталкивался на несообразности иерархического подхода, то отсюда он и сделал заключение об искусственности всякой системы вообще. Подходя диалектически, можно сказать, что, конечно, всякая наша система будет лишь приближением к объективной системе и каждый новый шаг будет все более естественным, мы все менее будем накладывать свои схемы на природу и все более приближаться к тому, что имеет место в природе. Это и будет синтез крайних мнений - кантианского (что мы законы природы налагаем на природу) и широко распространенного среди естествоиспытателей мнения, что найденные нами законы являются окончательными.
Можно вкратце противопоставить особенности иерархической системы в ее господствующем понимании и те особенности объективной системы, которые выясняются из размышления над конкретными системами. Для иерархической системы принимается:
В противоположность этому мы можем утверждать:
Основное: отождествление систематики и филогении, исторический подход к систематике. Это основание, конечно, теснейшим образом связано с логическим основанием. Если рассматривать систему только как отражение истории, не подчиненную своим закономерностям (антиномогенетическая точка зрения), то, естественно, покажутся удивительными все те расхождения с иерархической системой, о которых уже было указано. Говоря коротко, с филогенетической точки зрения сходство есть мерило родства, и сходство, основанное не на родстве, составляет небольшое число случаев среди сходства вообще.
Параллелизм, конвергенция и т. д., обилие случаев, где группы, считавшиеся монофилетическими, оказываются полифилетическими, все больше нас убеждают в том, что большинство сходств не имеет филогенетического значения и, говоря о системе как основанной на сходстве (естественная система), а не на родстве, следует по крайней мере усомниться в том, что система есть отражение филогении. Дарвин в последней главе "Происхождения видов" говорит, что системы, подобные биологической, имеют место и в других областях, например в химии, но там им нельзя дать того толкования, которое дается им для биологической систематики (филогенетическое), и он не знает, какое толкование может быть дано. За это время положение существенно изменилось: в химии и в других областях система получила рациональное объяснение, а в биологии то, что казалось Дарвину ясным, стало неясным.
Они заключаются в крайней отсталости методологии современной систематики в целом, хотя имеется большое число разрозненных и не связанных друг с другом попыток усовершенствования этой методики. Перечислю основные дефекты.
А. Крайняя неточность описания наряду с раздутостью - в этом отличие от кратких и достаточно точных для своего времени диагнозов Линнея. То, что было допустимо во времена Линнея при малом количестве известных видов, достаточно резко отграниченных, то совершенно недопустимо в настоящее время, когда число видов чрезвычайно увеличилось, а границы между ними стали гораздо более неопределенными. Читая описания видов, занимающих (например, в "Фауне СССР") обычно полстраницы, а иногда и целую страницу, видишь, что, кроме нескольких немногих фраз, все остальное - совершенно неопределенные высказывания, которые ни в какой степени не помогают определению. Бесполезность таких расплывчатых описаний сознают многие зоологи. Например, Гейкертингер (F. Heikertinger) прямо пишет, что определить виды группы Haltica oleracea без исследования копулятивного аппарата самцов невозможно. (То же касается многих видов рода Longitarsus.) Однако в сводках по систематике приводятся длиннейшие (и совершенно бесполезные) описания видов этой группы, как будто они могут чем-либо помочь. Мало того, эти описания при своей бесполезности иногда ухитряются вводить в заблуждение. Возьмем, например, описание Haltica carduorum и Н. oleracea в такой, вообще говоря, прекрасной (и до сего времени для этой группы непревзойденной) сводке, как книга Вейзе (J. Weise). Для Н. oleracea указывается, что ширина переднегруди в полтоpa раза больше длины, а для Н. cardliorurn-в два раза.
Различие как будто достаточно ясное. И если глазомером оценить отношение длины к ширине, сравнивая серии обоих видов, то получается ясное впечатление, что Вейзе прав и что этот признак действительно разграничивает два вида. Однако измерение с помощью микрометра показало, что впечатление основано на оптическом обмане. Измеряя по 20 экземпляров каждого вида, я нашел, что для Н. carduorurn это отношение равно 1,53, а для Н. Oleracea - 1,48, причем разница все же статистически реальна. Описание, таким образом, неверно. Оптический же обман объясняется, по-видимому, различным положением поперечной бороздки на переднегруди. У Н. olerecea она расположена значительно более кпереди, чем у Н. carduolurn (признак, как и все остальные, трансгрессивный, но на целой серии ясно выраженный). И это различное положение поперечной бороздки и создает впечатление большей вытянутости переднеспинки у Н. oleracea.
Другой пример - переднегрудь у Longitarsus exoletus. Во всех описаниях она обозначается как квадратная, т. е. длина и ширина одинаковы, но это, безусловно, неверно, и только по сравнению с другими видами она больше приближается к квадрату. Поэтому совершенно точное следование тексту определенных таблиц делает невозможным определение L. exoletus. Обычно, проверив по копулятивным органам, проведя сравнение с западноевропейскими экземплярами и убедившись, что налицо действительно L. exoletus, энтомолог начинает понимать, что слово "квадратный" обозначает: уклоняющийся от равенства продольного и поперечного размеров меньше, чем у других видов.
Наконец, приведу еще пример: Н. brevicollis относится к категории крупных Haltica (указывается примерно 4-5 мм). В группе видов Н. oleracea (3-4 мм) не указываются различия размеров для видов oleracea, carduorum, palustris, sandini, carinthiaca, и только для вида pusilla отмечается несколько меньшая величина. Таким образом, из существующих описаний нельзя усмотреть наличия трансгрессии между Н. brevicollis и oleracea и также нет и намека на наличие видовых различий по величине в пределах группы Н. oleracea.
Мое первое же измерение указанных видов показало сразу ошибочность обоих положений. Я не проделывал измерений больших серий oleracea и brevicollis, но собранный и оставшийся в Киеве обширный материал показал, что, например, в окрестностях Киева очень много мелких Н. brevicollis. Курьезно, что самый маленький самец Н. brevicollis оказался меньше самого маленького самца среди моей довольно обширной (свыше 200 экз. самцов) серии самцов Н. oleracea. С другой стороны, Н. oleracea имеет размеры существенно меньшие, чем Н. carduorurn и Н. palustris, хотя, конечно, этот признак является сильно трансгрессивным. Мы видим, таким образом, что там, где по описаниям вовсе нет трансгрессии, наблюдается абсолютная трансгрессия (амплитуда одного вида охватывает полностью амплитуду другого вида), а там, где не указывается никаких отличий, имеются отчетливо выраженные отличия, хотя бы и трансгрессивные. Несомненно, подобных примеров можно привести сколько угодно, и все они указывают, какой обширный резерв нетронутых признаков еще имеется в любой группе; этот резерв может быть использован при тщательном описании.
Все это объясняется тем, что в систематике до сих пор господствуют примитивнейшие методы описания. Размеры показываются наименьшие и наибольшие, т. е. применяется давно уже отвергнутая в биометрии амплитуда. Вполне естественно, что, так как амплитуда оценивается только на глаз, эта амплитуда оказывается различной у разных авторов. Если мы возьмем, например, два последних советских определителя (жуков и других насекомых), где таблицы по листоедам составлялись лучшими специалистами по этому семейству - Г.Г. Якобсоном и Д.А. Оглоблиным7, и сравним средние размеры (середины амплитуд) для девяти видов блошек рода Phyllotreta, то оказывается, что во всех девяти случаях размеры в работе Оглоблина показаны большими, чем у Якобсона, и средняя разница равна 0,328 мм при средней ошибке 0,067 мм. Различие настолько велико, что оно не могло бы возникнуть случайно, даже один раз на 1000 случаев. Если бы эти книги составлялись по материалам из разных стран или разных времен, то будущий исследователь, доверяясь указаниям лучших специалистов своего времени, мог бы сделать совершенно неверный вывод о параллельной географической или вековой изменчивости.
Ясно, что при описании нужно применять современные биометрические средства.
Б. Игнорирование количественных признаков и габитуса. Этот дефект, как и предыдущий, связан с неумением и нежеланием систематиков (конечно, не всех, но огромного большинства) пользоваться биометрией. Иногда под это подводятся и теоретические основания. Помню, как на одном съезде покойный П.П. Сушкин после доклада Ю.М. Вермеля говорил о недопустимости чисто количественных признаков. Правда, когда я заявил: "А аллометрон Осборна?"8, то он смутился и стал говорить что-то неясное. Но нет надобности призывать авторитет Осборна, чтобы показать, что дело объясняется просто косностью систематиков. Для отличия таких хороших родов, как Aphtona и Longitarsus, принимается обычно чисто количественный признак (первый членик задней лапки Longitarsus длиннее половины голени), но количественные признаки применяются в систематике только тогда, когда они бросаются в глаза настолько резко, что не требуют измерения. Между тем каждому систематику известно, что наряду с излюбленными в систематике признаками (наличие бороздок, форма частей тела, их взаимное расположение, число шипиков, члеников усиков и т. д.), которые у мелких видов могут быть видимы только при увеличении, имеются резко выраженные габитуальные отличия; даже мелкие виды родов Aphtona и Longitarsus можно отличить простым глазом, и не только знакомые виды, но и тогда, когда впервые попадает экземпляр незнакомого вида. Ясно, что есть какие-то четкие различия в пропорциях тела и эти различия можно использовать для характеристики трудных для определения родов и видов.
В. Недостаточная изученность сравнительной ценности разных признаков. Нельзя сказать, чтобы этот вопрос мало обсуждался: литературных высказываний и полемики было сколько угодно, но большей частью вопрос разрешается догматически, причем господствует страшная путаница и непоследовательность. Приведу пример. Во введении к "Флоре СССР" высказывается как непреложная истина (это как будто было принято на одном ботаническом съезде) следующее: следует твердо помнить, что между видовыми и родовыми признаками существует принципиальная разница, и, в частности, насколько мне помнится, для характеристики семейства берутся половые органы цветка. Это же повторяется и в книге "Учение о виде у растений" В.Л. Комарова9. Это отчетливо линнеевское понимание сравнительной ценности признаков (в частности, Линней совершенно отрицал сколько-нибудь значительную систематическую ценность окраски цветков) вполне гармонирует со взглядами Агассица10 (в книге "О виде и классификации в зоологии", 1869), утверждавшего, что даже если бы существовал всего один вид, то и тогда в нем можно было бы различать признаки типа, класса, отряда, семейства и т. д. У зоологов, напротив, господствует представление о чисто относительном характере признаков: систематическая ценность признака определяется чисто формально, а именно числом коннотаций (выражение Гельмгольца), связывающих этот признак с другими. Курьезнее всего то, что Комаров и другие, высказывающие подобные суждения, считают себя дарвинистами - явное отсутствие строгости в мышлении. Такое длительное переживание систематических идей Линнея и Агассица уже само по себе говорит, что в них, вероятно, есть какое-то здоровое зерно. Произвести синтез здорового начала во взглядах додарвиновских ученых и Дарвина - одна из задач общей систематики.
Другая задача: выяснить причины того значения, которое имеют копулятивные органы в систематике. Совершенно ясно, что небольшие изменения в скульптуре копулятивных органов, например Haltica и Longitarsus, не носят приспособительного значения, но остается непонятным, почему копулятивные органы так часто оказывают ценные услуги систематике. Этот вопрос, конечно, недостаточно изучен. Мы знаем, что есть виды, хорошо отличимые по соматическим признакам и не дающие резких отличий по копулятивным органам (видимо, например, большинство видов рода Chaetocnema и также многие Phyllotreta), и, напротив, есть виды, резко отличимые по копулятивным органам и почти неотличимые по соматическим признакам (многие На1tica и Longitarsus). Если изменения копулятивных органов независимы от изменений соматических, то должны существовать и две остальные комбинации: виды, хорошо отличимые и соматически, и по копулятивным органам, и, напротив, "хорошие виды", почти не отличимые ни соматически, ни по копулятивным органам. Необходимо подвергнуть основательной переработке, например, группы Halticinae и Galerucinae, сравнивая изменчивость ряда признаков соматических и копулятивных. По-видимому, действительно четвертая категория существует, но, конечно, ее существование очень трудно доказуемо и просто невозможно обычным путем, так как у подобных видов нет отчетливых различий ни в соме, ни в половой сфере. Одной из первых работ в этом направлении надо считать работу Матер и Добржанского11 о чисто генетических расах Drosophila pseudoobscura, ведущих себя физиологически как "хорошие виды" (не дают плодовитого потомства), но при применении тонких биометрических приемов обнаруживающих и морфологические отличия. Из моей практики могу привести такой случай: весьма возможно, что Haltica carduorurn из Средней России и Черноморского побережья Кавказа-два вида, а не один. Я это заключаю по следующим соображениям.
Если окажется (а первое впечатление, что именно это имеет место), что копулятивные органы, так сказать, приходят на помощь систематику преимущественно там, где различий соматических признаков недостаточно, т. е. что есть известная отрицательная корреляция между размахом изменчивости соматических и копулятивных органов, то это составит крупную проблему.
Разрешение ее мыслимо, например, в следующих двух плоскостях.
Новая систематика (не в смысле "новой систематики" Дж. Хаксли13, где очень мало и нового, и систематического) уже показывает ряд свежих ростков. Отмеченные недостатки уже вызвали к жизни ряд попыток реформы систематики или искание новых путей. Но в большинстве эти попытки являются разрозненными, отдельные авторы не знают друг друга, некоторые математические работы остаются неизвестными (или неиспользованными) не только биологам, но и математикам. Перечислю главнейшие направления, причем, конечно, мой перечень будет крайне неполным, так как многие работы попадаются совершенно случайно и мне приходится сейчас цитировать только по памяти.
Попытки построить филогению в пределах рода или семейства
Эти попытки заключаются в том, что, исходя из представления о естественной системе как филогении, пытаются построить конкретную филогению группы, сравнивая число сходств и различий в пределах сравниваемых групп. Таких попыток мне известно три, вероятно, на самом деле их гораздо больше. Это работы Эйзиг в монографии о капителидах, Коллери по червям рода Spirorbis и Д.А. Смирнова о долгоносиках рода Pliyllobius14. Все эти попытки интересны тем, что построить филогению путем учета числа сходств и отличий не удалось и потому ценность этих работ скорее критическая, чем положительная. Но несомненно, этот путь можно развить, если, сохранив сам мeтoд исследования, отказаться от поставленной указанными авторами цели - уложить во что бы то ни стало систему в прокрустово ложе филогении, а рассматривать филогению как совершенно самостоятельную дисциплину. Весьма возможно, что, продумав основательно характер признаков (первичный, вторичный), можно прийти к заключению в отношении ряда признаков об их заведомо полифилетическом происхождении. Весьма возможно, что для ряда признаков удастся показать неизбежное принятие их обратимости и тем самым показать, что по крайней мере в определенном, не слишком широком интервале закон Долло неприменим даже в самом узком его понимании (невозможность восстановления утраченного органа). Правда, сейчас уже введено много оговорок к этому закону: поправка П.П. Сушкина15 о возможности обратимости путем неотении; появление исчезнувшего пальца, кажется, у полевок (по Б.М. Житкову, из В.М. Шимкевича); не слишком редкое появление крылатых особей у видов, утративших крылья. Здесь, конечно, как и всегда, надо быть осмотрительным: мне казалось таким ярким примером обратимости наличие летающих карабусов (статья в Koleopterologische Rundschau), но, по указанию Арнольди, это вовсе не возврат, а просто у многих видов рода Carabus имеются крылатые разновидности, которые даже доминируют в некоторых местностях. Видимо, закон Долло имеет место только тогда, когда по линии редукции какого-либо органа проделана сложная эволюция, если же орган исчез быстро, то этот процесс вполне обратим. Этот вывод, по-видимому, можно тщательно обосновать, именно следуя по пути указанных авторов, и тогда станет совершенно ясно, что актуальное развитие данного признака имеет в основном систематический, но отнюдь не филогенетический смысл. Последний он может приобрести только на значительных систематических интервалах. Эти попытки имеют поэтому не только критическое (отрицающее) значение (это значение может быть поставлено в параллель с негативными выводами в математике о невозможности трисекции угла, о невозможности решения в радикалах уравнений выше четвертой степени и т.д.), но вместе с тем служит толчком для второго направления в систематике, пока еще очень робкого, относительно возможности построения иных форм системы, кроме иерархической.
Эти попытки можно проследить далеко в глубь веков - они делались еще до торжества эволюционной теории. Дарвин цитирует закон аналогичной изменчивости Уолша (в "Происхождении видов"), П.П. Сушкин мне указывал, что в первой половине XIX в. были работы Каупца, который в каждой систематической группе (птиц) устанавливал пять членов и, в частности, в каждой такой группе принимал один пестрый и один крупный вид. П.П. Сушкин говорил, что очень часто получалось хорошее совпадение. Литература по этому вопросу чрезвычайно обширна: закон гомологических рядов Н.И. Вавилова, периодическая система пантопод В.М. Шимкевича и мн. др. В частности, в одной из работ Гейкертингера16 о Psilliodes виды распределены в такую решетку. Нетолицкий применял этот же принцип для определительных таблиц рода Bembidium. Есть одна работа, автора которой сейчас не помню (ее указал мне Д.Е. Харитонов в Перми), где автор пытался по степени сходств определить расстояние между видами и затем построил чертеж, так сказать проекцию, естественной системы с указанием расстояний между видами. Все это, конечно, только первые попытки, так как решетчатое строение системы (о сети, как об отображении системы говорили в свое время К. Линней и И. Жоффруа Сент-Илер) является только первым шагом по пути отхода от иерархической системы и по пути продвижения к подлинно естественной системе.
Использование комплексных признаков.
Это направление уже касается не значительных систематических категорий, а, напротив, мелких систематических единиц и свое начало ведет от Гейнке17, применившего объединение нескольких трансгрессивных признаков для различения рас сельдей. В самой работе Гейнке и материал невелик, и обработка довольно примитивна, есть, кроме того, ряд совершенно необоснованных выводов, например о том, что сумма квадратов отклонений каждой особи от средних по ряду признаков есть величина постоянная для расы (это положение необоснованно, но, может быть, заключает долю истины в связи с постулатом о константе формообразовательной энергии). Кроме того, метод Гейнке в его первоначальной форме страдает тем недостатком, что он пригоден только при полной независимости использованных признаков, в случае же коррелятивной зависимости могут получиться грубые ошибки (что и случилось как будто в Средней Азии при применении к Homalocopris tmolus). В силу всего этого попытки применений метода Гейнке немногочисленны и больше всего их как будто в русской литературе: в работе Ю.А. Филипченко18 о хермесах (кстати сказать, ни эта работа, ни работа Гейнке не цитируются в одной довольно обстоятельной сводке о биологических видах одного американского автора, которую я видел у Блисса - пережевывание пустой болтовни о биологических видах), П.В. Серебровского в работе о курах, некоторых ихтиологических работах и в неопубликованной работе С.С. Четверикова о стрекозах.
Путь к преодолению основного дефекта метода Гейнке (применимость только в случае независимых признаков) показал В. И. Романовский в одной статье в "Трудах Среднеазиатского Университета"19. Эта статья элементарно простая, но чисто методическая, без конкретного материала. Он указал, что при применении корреляционной таблицы по двум признакам при сильной трансгрессии обоих признаков может быть случай, когда эта трансгрессия в двухмерной области исчезает.
Поместив экземпляры обоих видов в виде точек на плоскость, мы видим, что, рассматривая каждый признак сам но себе и беря его амплитуду (хотя бы в виде трех сигм в обе стороны от арифметической средней), мы получаем для каждого признака в отдельности очень сильную трансгрессию. Но как только мы обратим внимание на эллипсы рассеяния обоих видов и ограничим эллипсы прямыми параллельными линиями (хотя бы в виде трех сигм не от арифметического среднего, а от линии регрессии), так тотчас же трансгрессия исчезает.
От этой работы логически (но не хронологически и не генетически, так как работы Эджвурт-Пирсона были задолго до работ Романовского, а Р. Фишер, видимо, не знает работ ни Гейнке, ни Романовского) ведут начало три направления.
А. Непосредственное продолжение метода Гейнке - Романовского, которое я провел в сданной в печать в Киеве работе о различии личинок Melolontha melolontha и М. hippocastani. Вместо расстояния величины данного признака от средней арифметической, деленной на сигму, я взял расстояние значения данного признака от линии регрессии по другому признаку, деленное на среднюю сигму от линии регрессии, и квадраты этих величин суммировал, как это делал Гейнке для 5-7 признаков. В результате в большинстве случаев можно было различать виды, причем в главной массе совершенно надежно: я сделал попытку, довольно грубую, оценить значимость получаемых сумм квадратов. О применении номограмм в данном случае речь будет дальше.
Б. Усовершенствование метода комплексных признаков. Здесь нужно отметить работу Р. Фишера20 (Annals of eugenics, около 1936 г.), не связанную ни с первым, ни с третьим направлением. Он ставит задачу так: положим, два вида различаются по ряду признаков. Каждую функцию (он берет для иллюстрации только линейную функцию четырех признаков) от этих признаков надо взять, чтобы различие между видами было максимальным. Он решает эту задачу, применяя свой дисперсионный анализ. Он приводит все вычисления для избранного им объекта трех видов рода Iris. Однако с биологической точки зрения объекты им выбраны очень неудачно, так как почти во всех случаях не наблюдается никакой трансгрессии. Поэтому такая сложная аппаратура для решения очень просто решаемой задачи может вызвать справедливую реплику: "веревка-вервие простое"... Этим же методом пользуются Матер и Добржанский (см. выше) в своей очень интересной работе о Drosophila pseudoobscura, но там они допускают противоположный дефект: берут несколько признаков и механически применяют к ним методику Р. Фишера, не замечая, что из примененных ими признаков некоторые не показывают и намека на существенные различия и что поэтому их включение не приносит никакой пользы для диагностики (что и оказалось по окончательным таблицам), значительно затрудняя вычисления. Необходимо этот тонкий метод применять к признакам, отобранным на основе рекогносцировочного анализа.
Для меня работа Р. Фишера представляет интерес. При дальнейшей работе по замене определительных таблиц номограммами придется, естественно, ограничивать число шкал: вряд ли можно на одной номограмме практически пользоваться более чем 4-6 признаками. Но если не удается подобрать такие 4-6 признаков, которые ликвидировали бы трансгрессию, то естественный выход-заменять одиночные признаки комплексными признаками в смысле Р. Фишера и этими новыми признаками пользоваться при конструкции. Тогда потребовалось бы для 16 признаков пять номограмм-одна основная и четыре дополнительных.
В. Теорема Эджвурта-Пирсона о вероятности принадлежности данного индивида к определенному виду или вообще группе. Теорема Эджвурта-Пирсона изложена, например, в книге Слуцкого21 о теории корреляции (1912) и является обобщением гауссовой кривой. В показателе е-функции Эджвурта - Пирсона стоят не только сумма квадратов нормированных отклонений нескольких признаков от соответствующих арифметических средних (в этой части функция соответствует произведению вероятностей нескольких независимых событий - первоначальный метод Гейнке), но и нормированные коварианты по всем парам признаков. Мне не известно ни одной работы, где бы эта функция применялась к конкретному биологическому материалу, хотя, например, Е.С. Смирнов ее цитирует. Видимо, комбинируя использование этой функции с моим методом использования метода Гейнке-Романовского, можно получить более точную оценку надежности определений, чем при помощи того грубого приема, который я применял в работе о личинках Melolontha. Вероятно, эта функция разобрана в книге Романовского "Математическая статистика"22.
Перейду теперь к краткому изложению основных разделов общей систематики.
ПРАКТИЧЕСКАЯ ОБЩАЯ СИСТЕМАТИКА
Она состоит из двух частей: аналитическая, или идиографическая, систематика; синтетическая, или номографическая, сиcтематика.
Аналитическая систематика - идиография
В область идиографии23 входят три задачи:
1. Описание. Хотя в настоящем разделе мы касаемся практической систематики и задача описания как будто сужена практическими потребностями, не следует понимать практику в смысле голого определения. Слово "практический" имеет общеметодический смысл, и потому вопрос об описании надо поставить во всей широте, и прежде всего поставить вопрос: возможно ли полное описание, т. е. возможно ли охватить конечным число слов все богатство признаков морфологических, эмбриологических, физиологических, химических и т. д. так как все такие признаки могут быть использованы как для самой систематики так и для других целей. Обычный взгляд на организм как на агрегат независимо изменяющихся частей и признаков, конечно не допускает возможности действительно полного описания. С этим взглядом стоит в связи и чисто генетическая трактовка системы, и такое частное мнение, как допущение существования биологических видов и рас, т. е. таких, которые различаются исключительно физиологическими, экологическими и другими признаками при полном сходстве морфологических признаков. Напротив, взгляд Скиапарелли на организм как на чистую форму (т. е. такую, по любому, сколь угодно малому отрезку которой можно восстановить всю форму) даже в модифицированном отражении (принимая организм как комплекс чистых форм) допускает возможность полного описания. Поэтому сейчас ряд биологов выдвигают положение, что, может быть, число истинных признаков организма не бесконечно, а конечно. Этот взгляд получил свое выражение, например, в одной из работ П.В. Терентьева, который разделяет все возможные плеяды, связанные коррелятивной связью, и тогда достаточно будет от каждой плеяды знать один или несколько ведущих признаков, чтобы на основании их сконструировать весь организм. Не пытаясь решить этот вопрос (для этого необходимо проделать еще очень много конкретных исследований), можно сказать, что, во всяком случае, надо стремиться к тому, чтобы связывать ряды признаков в системы коррелятивных связей, и тогда описание приобретет гораздо большую полноту при гораздо большей краткости. Под ведущими признаками следует понимать такие, которые всего удобнее принимать как независимые при определении по ним иных зависимых признаков. При точном описании в отношении каждого признака надо определить не только его среднюю величину (арифметическую, геометрическую, медиану, моду), но и дисперсию, в общем случае построить кривую распределения. Вероятно, можно выставить такой биологический постулат, что, как бы ни были близки между собой две таксономические группы, нельзя найти ни одного признака, по которому они не отличались бы. Например, все млекопитающие имеют четыре конечности, лишь китообразные имеют редуцированные задние конечности. Не говоря уже о том, что в последнем случае, введя балл для характеристики развития задних конечностей, можно характеризовать животное дробным числом конечностей; даже в том случае, когда конечности имеют полное развитие, нельзя говорить о полном тождестве их числа для любых двух групп организмов; бывают млекопитающие уроды с редуцированным числом конечностей (описана даже родившаяся совершенно безногая овца). Склонность к таким уродствам различна у разных видов, и, беря генеральную совокупность особей двух соседних видов и изучая кривые распределения, мы, весьма возможно, найдем различие и по этому, казалось бы, совершенно стабильному признаку.
Очень важное понятие - кондициональные признаки (о них говорится и у Дарвина), на что обращал внимание В.Н. Беклемишев в лекциях по теории системы в Пермском университете. Группа может характеризоваться не тем или иным признаком, выраженным у всех своих индивидов, а определенной способностью проявлять тот или иной признак при каких-то, нам очень часто вовсе неизвестных, условиях. Хороший пример таких кондициональных признаков - образование вдавления на брюшке у самцов ряда родов Gallerucinae (Phyllobrotica, Euluperus, Luperus; см. монографию Оглоблина в "Фауне СССР", с. 189, 191, 195, 201, 205, 279, 281). В разных родах это вдавление, несомненно, развивалось независимо, что можно заключить как из серии постепенного развития таких вдавлений в каждом роде, так и из отличительных родовых черт таких вдавлений. Кондициональным признаком для таких групп, как Orihoptera (Saltatoria), является способность образовывать совершенно между собой несходные стридуляционные органы; то же - стридуляционные органы совершенно различного характера-наблюдается у личинок в ряде родов пластинчатоусых жуков. Упомяну еще любопытный факт (видел целый атлас южноамериканских гусениц сфингид в Nature novitates, издание музея Ротшильда) - образование рисунка, напоминающего голову змеи на самых разнообразных местах тела (передний, задний конец и т. д.) гусениц сфингид. Можно указать и сравнительно редкий случай, где мы можем выставить и причину реализации кондициональното признака: при переходе членистоногих от водного образа жизни к наземному могут образоваться очень сходные органы: трахеи насекомых, сольпуг и мокриц.
Весьма возможно, что при дальнейшем углубленном изучении в связи с торжеством идеи о полифилетическом происхождении разных групп большинство признаков окажутся кондициональными. В пользу этого говорит уже одно то, что в самых естественных группах (что известно было и Дарвину) очень трудно найти признаки, характеризующие данную группу в целом, например ракообразные. Много примеров в отношении систематики блошек у Гейкертингера; из чтения книги С.А. Северцова24 "Динамика населения..." (1941) видно, что, например, низшие сумчатые вообще сумки не имели, а у высших она построена очень разнообразно. Плацента имеет у плацентарных очень разнообразное строение. Эти, видимо, общепризнанные факты позволяют уже утверждать, что плацентарные, сумчатые и т. д. характеризуются вовсе не плацентой, сумкой и т. д., а лишь способностью произвести сумку, плаценту и т. д. Причем эта сумка или плацента вовсе не предопределена как таковая, а лишь в форме потенции, могущей быть воплощенной самым разнообразным образом. Можно пойти еще дальше. Плацентой обладают не только млекопитающие, но и некоторые рептилии, а живородящие свинки тупохвосты (Trachysaurus rugosus) обладают настоящей аллантоидной плацентой (Пузанов, 193825, с. 328, см. 121, 5 / Цифры 121.5 означают том 121 архива, позиция 5. (Примеч. ред.)). Мне помнится, что плацента имеется и у некоторых акул, скатов и у некоторых живородящих насекомых. Любопытно было бы сравнить плаценты в столь широком сравнительно-анатомическом разрезе. С точки зрения кондициональных признаков весьма вероятно, что между некоторыми формами плаценты млекопитающих, акул, рептилий и пр. окажется больше сходства, чем между плацентами в пределах одного класса. В пользу того, что большинство признаков, характерных, например, для млекопитающих, окажутся кондициональными, говорит одно примечание в той же книге Северцова о динамике населения. Он пишет: или надо принять самостоятельное развитие важнейших признаков млекопитающих (волосы, млечные железы и т. п.) у разных групп млекопитающих, или допустить, что эти признаки были свойственны очень древней группе. Видимо, последняя альтернатива принимается эпигонами дарвинизма только в силу косности мышления.
Наконец, можно привести еще один прекрасный пример кондициональных признаков: расширение члеников передних лапок у самцов жуков. В самых разнообразных семействах (Carabidae, Tenebrionidae, Dytiscinae, Halticinae и т. д.) мы имеем роды, где самцы имеют расширенные лапки (причем с разным числом члеников), и другие роды, где расширения нет. Ясно, что это расширение произошло независимо в разных семействах и потому, может быть, для всех жуков кондициональным признаком является расширение члеников передних лапок.
Кондициональные признаки не могут рассматриваться как простое осуществление скрытого зачатка. Против этого говорит чрезвычайное многообразие проявлений кондиционального признака. Напротив, их скорее можно сравнить как бы с картиной, отброшенной проекционным фонарем на колеблющийся экран. Наилучшую аналогию этому можно найти в диалоге Платона "Государство" (в пещере на стене отражаются тени проходящих мимо людей). Идеи Платона могут иметь еще эвристическое значение и быть использованы для осмысления актов, примитивно толкуемых эпигонами дарвинизма (например, "девиации" А.Н. Северщова).
Понятие кондициональных признаков в значительной мере стирает границу между гомологичными и аналогичными органами. Я лично склонен думать, что понятие гомологии не носит обязательно генетического характера в смысле преемственности актуализированных форм, но может быть преемственностью в той потенциальной форме, динамическом эмбриональном поле А.Г. Гурвича26 и т. д., которую можно назвать гомологией высшего порядка. Само собой разумеется, что ведущие признаки нам придется отбирать из совокупности реальных, а не потенциальных признаков и этот путь отбора заключает в себе много исканий. Путь, который я проделал с личинками Melolontha и начал с видами рода Haltica, заключается в том, что для нескольких десятков признаков производится сравнение средних и их дисперсий. Здесь мы переходим к проблеме разграничения видов и других таксономических единиц.
2. Разграничение. Для простого описания ценность признака - в тесноте его корреляционных связей с другими признаками (обилие коннотаций по Гельмгольцу). Для разграничения соседних таксономических единиц выдвигаются другие критерии. Здесь, как я уже указывал, всплывают вопросы о сравнительной ценности и иерархической подчиненности разных признаков. Эта сравнительная ценность может определяться разными критериями и вряд ли может быть выставлена a priori.
Чисто формальные критерии: первый - отсутствие или минимальная величина трансгрессии. В работе с Melolontha я применял очень простой критерий, названный мною коэффициентом дивергенции (в литературе этот термин применялся в другом смысле) и выражающийся формулой
D=(M1-M2)2/(s12+s22)
(квадрат разности медий, отнесенный к сумме дисперсий, квадратов сигм). Мы можем принять, что трансгрессия отсутствует (для простоты примем, что сигмы обоих видов равны), если расстояние между средними арифметическими равно шести сигмам (т. е. отклонение плюс три сигмы от одной средней как раз совпадает с отклонением минус три сигмы от другой). В таком случае этот коэффициент, очевидно, равен 18. В моей работе о личинках Melolontha "лучшие" признаки показывали коэффициент дивергенции, равный 0,8, и даже при таком малом коэффициенте комбинация нескольких признаков позволяла определять виды в большинстве случаев. Для ориентировочного определения реальности различий можно пользоваться таким упрощенным приемом: располагают исследуемый признак в восходящий и нисходящий ряды обоих видов, затем считают число экземпляров обоих видов в области трансгрессии и относят его к общему числу экземпляров. Если процент (при общем количестве в несколько десятков) меньше 50, то можно уже с уверенностью говорить о реальности различия. Из взятых для испытания ряда признаков отбирают те, которые показывают наибольшие коэффициенты дивергенции. Эти уже отобранные признаки затем сводятся в комплексы по методам, намеченным Романовским и Фишером.
Второй формальный критерий - зависимость от экологических и географических различий. Эта зависимость имеет и недостатки, и достоинства. Признаки, зависящие от экологических и географических различий, конечно, мало пригодны для характеристики видов в целом, так как они увеличивают общевидовую дисперсию, но зато могут быть прекрасно использованы для определения географического положения и характеристики низших таксономических единиц.
Что же касается сравнительной значимости признаков, то по этому вопросу литература очень обширна, но обоснование тех или иных положений или очень туманно, или вовсе отсутствует. Например, человекообразные обезьяны имеют те же группы крови, что и человек, и, видимо, можно переливать кровь от обезьян человеку в пределах той же группы. По этому признаку того же "кровного" родства выйдет, что надо образовывать родственные группы, в каждую из которых войдут некоторые люди и некоторые обезьяны. Такую уродливую форму полифилетизма, я полагаю, никто защищать не станет, С другой стороны, можно привести ряд примеров, где правильная морфологическая группировка влечет за собой неизбежно и соответствующую биохимическую. Особенно показателен случай с комарами Culex и Anopheles. Различие родов было установлено, когда еще не была известна связь последнего рода с малярией, и, однако, изучение малярии вполне оправдало чисто морфологическую систематику. В противоположность биохимическим признакам, которым придается чрезмерно большое значение, есть признаки, которым отказывают в каком бы то ни было значении (например, окраска цветков по Линнею). Помню, А.П. Семенов-Тян-Шанский восставал против аналогичных утверждений энтомологов отказывать в таксономической ценности окраске. Он указывал, что есть такие группы, где окраска имеет определенное таксономическое значение. Даже у растений можно указать, например, на водоросли - бурые, зеленые, синезеленые, красные, где цвет является ведущим признаком крупных систематических единиц27. Но в общем Линней, конечно, прав: такие случаи, где цвет играет важную таксономическую роль - редкое исключение. На данном примере, пожалуй, мы действительно видим сохранение идей Линнея или Агассица о том, что определенным признакам свойствен определенный таксономический уровень. Копулятивные органы, по мнению многих, имеют особенно высокое систематическое значение, но неодинаковое таксономическое положение. Многие им приписывают особое значение для разграничения видов (в связи с общепринятым, хотя и не вполне правильным положением о бесплодии гибридов между видами), но не придают большого значения для различия высших систематических категорий (в ботанике имеет место другое, как уже было указано). Однако мы имеем и обратные случаи. Н.А. Теленга, например для браконид, выдвигал, что копулятивные органы очень важны для различения родов, но мало пригодны для различения видов. Для Halticinae, например, их пока применяли только для различения видов, и мой собственный опыт по ряду родов показывает, что при огромном разнообразии эдеагусов в пределах некоторых родов пока не представляется возможным выделить какие-то родовые признаки для копулятивного аппарата. Мало того, в строении эдеагусов мы наблюдаем конвергенцию не только в пределах рода (Longitarslis nigrofasciallis, L. brisuti), но и между родами: с эдеагусом этих двух видов очень сходен, отличаясь в общем только шириной, эдеагус довольно отдаленного рода - у Chaetocnema semicoerulea. Получающийся вывод о том, что конвергенция копулятивных органов имеет примерно то же распространение, что и в пределах других систем, надо тщательно проверить и сопоставить с высказываниями авторов, стремящихся доказать, с одной стороны, что по строению эдеагусов или внутренних мешков пенисов можно хорошо построить иерархическую естественную систему, являющуюся вместе с тем филогенетической, а с другой - что имеет место тесная зависимость в развитии мужских и женских копулятивных органов (см., например: Даль - для пауков, Гольдхаус - для Microlestes из карабид, Жаннель - для Batiscinae28 и др. указания в одной из обзорных статей Гейкертингера в "Коleopterologische Rundschau" в конце двадцатых годов). Не исключена, таким образом, возможность, что, как правило, определенным ступеням системы будет соответствовать особая значимость различных признаков: окраска для некоторых категорий ниже вида, детали копулятивных органов для вида и т. д. Этим путем, может быть, удастся выделить истинное зерно в положении Агассица о таксономической неравноценности признаков.
Выявив сравнительную таксономическую ценность разных признаков на основании попыток "полного описания", можно будет приступить к решению задачи о дивергенции не только отдельных признаков, но и для генеральной совокупности признаков сравниваемых видов. Попытки в этом направлении уже делались, но как будто без достаточного продумывания самой возможности полного описания вида. Я помню статью Лахтина в "Антропологическом журнале" (в начале послереволюционных годов) о коэффициенте сходства. Особое внимание надо обратить на изучение пропорций тела и вообще на габитуальные признаки, находящиеся до сего времени в крайнем пренебрежении. Между тем несомненно, что они-то и руководят систематиками при разбиении на естественные роды и лишь потом подыскиваются морфологические признаки для большего удобства и для удовлетворения господствующей традиции. Конечно, вероятно, что роды отличаются друг от друга не только количественно, но и качественно в разных группах: например, весьма вероятно, что роды у ракообразных с морфологической точки зрения обширнее, чем у насекомых, у млекопитающих - обширнее, чем у птиц. Роды у Halticinae в большинстве случаев очень естественны, характеризуясь и габитусом, и структурными признаками, но роды, группирующиеся около Podagrica или Crepidodera, часто характеризуются более формально: этот формальный подход, может быть, особенно резко выражен в подсемействе Galerucinae в группе родов, близких к Galeruca. Получив критерий сходства в общем виде, можно подойти к вопросу о количественном и качественном различиях родов в различных семействах и т. д.
3. Таксономические единицы, разных порядков. По этому вопросу литература очень обширна, я же сам пока очень мало думал, так как, естественно, плодотворные мысли могут возникнуть лишь после значительной работы по оценке сравнительного объема и качественного различия разных систематических единиц. Для таксономических единиц ниже вида, конечно, надо проштудировать работу А. П. Семенова-Тян-Щанского, а для ознакомления со взглядами генетиков - сводки Добржанского и Турессона29.
Три единицы ниже вида довольно отчетливы: подвид (в основном определен географически), морфа (в основном определена экологически) и аберрация (определена исключительно генетически). Конечно, и подвид определен тоже генетически и при этом актуально, а морфа определена генетически кондиционально. (Здесь снова встречаемся с кондициональными признаками.) Очевидно, на очереди стоят вопросы о географической изменчивости видов, о сравнительном характере изменчивости разных таксономических единиц и т. д.
Синтетическая практическая (номографическая) систематика.
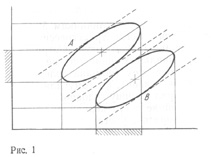
Простейшее применение мною номограмм к личинкам Melolontha показало, что они различаются (всегда трансгрессивно) по ряду признаков, из них два признака (число крючков стридуляционного аппарата и число преданальных шипиков) независимы один от другого и от остальных трех (ширина головной капсулы, длина 2-го и 4-го члеников усиков), поэтому для них и для одного из остальных трех (я брал ширину головной капсулы - c) можно применять метод Гейнке в его первоначальном виде, остальные же признаки надо брать с учетом их коррелятивной зависимости. Положим (рис. 1; линии регрессии, ограниченные полосами ±3, показаны сплошными и пунктирными линиями, а штриховкой отмечена область трансгрессии видов А и В), b - длина 2-го членика усиков, c - ширина головной капсулы. Тогда b - b=Ρ(c - c), где Ρ - коэффициент регрессии, а буква с черточкой обозначает среднее арифметическое данного признака. Квадрат относительного отклонения наблюденного значения от вычисленного будет равен
v=[(b - b) - (c - c)]2/(Σb)2
где Σb - стандартное отклонение b от линии регрессии. Извлекая корень, получим
g=[(b - b) - (c - c)]/Σb
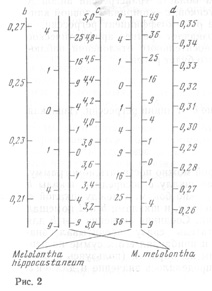
-линейная функция, для которой можно построить номограмму. Для экономии места я помещал шкалу c посередине, шкалы b и d (длина 4-го членика усика) - по бокам, а между шкалой c, принятой за независимое переменное, и шкалами b и d помещал шкалы обоих видов с отметками. Соединяя значения с и b для определенного экземпляра, читатель сразу прочитывал значение для данного экземпляра и прибавлял его к сумме по первым трем признакам. Подобным же образом (пользуясь правой частью номограммы; рис. 2) определялось значение и для d и c. Σv по пяти признакам и дает материал для суждения о принадлежности данной особи к тому или иному виду. Чем больше эта сумма, тем меньше вероятность принадлежности данного экземпляра к данному виду. В работе о Melolontha я пытался определить рассеяние эмпирически, но, мне думается, что если Σv получается от n признаков, то (Σv)1/2/n для своего вида колеблется около единицы со стандартным уклонением, равным 1/(n)1/2. Это надо проверить на материале по блошкам и посмотреть работы дельных математиков, может быть Р. Фишера.
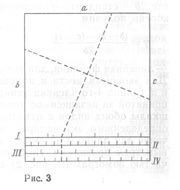
Конечно, такие номограммы могут быть построены и не для двух, а для нескольких видов. Кроме того, можно соединить в квадратной номограмме три независимых признака (рис. 3) и, пользуясь крестообразным транспаратном, прочитывать на четвертой стороне сразу сумму для трех признаков. Для оценки значимости рассеяния следует использовать функцию Эджвурта - Пирсона.
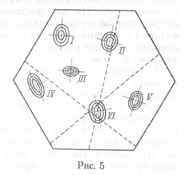
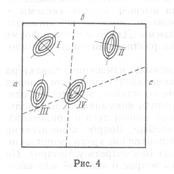
Я думаю развить метод номограмм в следующем направлении: положим, мы имеем четыре признака - a, b, c и d. Расположим значения их на четырех сторонах квадрата (рис. 4) и, соединяя попарно значения четырех признаков, по положению точки пересечения в поле того или иного вида судим о принадлежности данного экземпляра к тому или иному виду; на рис. 4 изображено поле для пяти видов. Концентрические эллипсы рассеяния соответствуют различным зонам значимости отклонения от центра вида. Степень надежности определения устанавливается по расстоянию точки пересечения от центра вида: чем меньше это расстояние, тем надежнее определение. Можно строить два квадрата для восьми признаков с целью контроля определения, а всего лучше построить гексагональную номограмму для трех пар признаков (рис. 5). В хорошо построенной номограмме три линии должны дать в пересечении маленький треугольник в поле одного из видов: если треугольник очень велик и если он не попадает в поле ни одного вида, то это значит, что или номограмма построена плохо, или мы имеем неизвестный вид. Из разговоров с М.Л. Франком, Л.Н. Бескиным и др. как будто выходит, что математики-номографы подобного рода номограмм не строили. Можно было бы помещать в каждую такую гексагональную номограмму лишь признаки одной плеяды (по терминологии П.В. Терентьева). Тогда три таких номограммы дали бы на трех страницах точное описание 18 признаков для целой серии видов: точность и сжатость описания чрезвычайно бы выиграли.
ТЕОРЕТИЧЕСКАЯ ОБЩАЯ СИСТЕМАТИКА
Здесь я приведу только краткие общие соображения, поскольку эта часть должна развиваться в тесной связи с развитием практической общей систематики. Теоретическая общая систематика распадается по крайней мере на четыре ветви:
Это - вопрос о форме системы и как таковой подвергся предварительному рассмотрению в моей печатной статье (1923) о форме естественной системы организмов. Она должна развиваться в тесной связи с номографией систематики, но номография возможна без номотетики и обратно: можно построить естественную систему, не отображая ее в номограммах. Между ним: даже большое существенное различие: номографическая систематика стремится удовлетвориться минимальным количеством признаков, номотетическая же должна охватить максимально их количество. Но для построения высшей формы системы - коррелятивной - чрезвычайно важно и полезно использовать материал, легший в основу номограммы. Для суждения о форме системы полезно размышление над различными видами рациональных систем.
Систематика и филогения, очевидно, совершенно различные дисциплины, но связь между ними несомненна. И этот вопрос заслуживает специального изучения. Возможно, например, что в естественной системе существуют линии преимущественного развития - то, что можно назвать геодезическими линиями системы. В пользу существования таких линий говорит частота параллелизмов в палеонтологии. Следует проработать для этого некоторые хорошо изученные филогении, например филогению лошадей, некоторых моллюсков, насекомых. Если выяснится, что изменение некоторых признаков имеет филогенетическое значение, то надо проанализировать эти признаки 1) с формальной стороны, в смысле характера их изменчивости, 2) с биологической стороны.
Проблемы, аналогичные связи между систематикой и филогенией, могут быть и в других науках: 1) в химии-периодическая система элементов и филогенез элементов (изотопы, полифилетическое происхождение компонентов, элементов); 2) в органической химии - система соединений и методы их получений; 3) в петрографии, как я уже указывал, видимо, есть такие горные породы, которые в силу разного происхождения попадают в разные места генетической системы, несмотря на тождественность свойств.
Здесь, очевидно, надо пересмотреть вопрос о логическом смысле разных таксономических единиц, сравнить с высказываниями формальной и индуктивной логики (Аристотель, Милль, Зигварт; посмотреть у Гегеля), постараться найти иную характеристику низших систематических единиц вместо ходячей "per genus proximus et per differentiam specificam". Кроме того, следует подойти к вопросу о природе различных таксономических единиц. Для этого очень полезно поразмыслить над природой таксономических единиц, например, в филателии (коллекции марок). Здесь очень отчетливо можно провести границу между видом и разновидностью. Вид - это идея марки, воплощенная художником в рисунке. Но филателисты отличают для каждого вида марок еще разновидности: по оттенку краски, по некоторым уклонениям клише, по числу зубчиков и т. д. Так как все эти отклонения не входили в идею марки, то все это единицы ниже вида. Но и в пределах их можно отличать качественно различные единицы: число зубчиков дает параллельные значения у марок различного достоинства. Это разновидность (подобно, например, форме аптера у многих насекомых). Уклонения в клише, вообще говоря, не повторяются у различных видов: это - настоящие аберрации. В филателии можно подыскать и таксономические группы выше вида: так все марки одного рисунка, но различной ценности и окраски, очевидно, образуют род. Подрод получается тогда, когда, положим, при одинаковом портрете или гербе имеется различное обрамление. Как и в систематике организмов, так и в систематике марок видовые различия в пределах рода могут быть различного диапазона, например: марки разной ценности могут (как это, например, имеет место в английских марках) иметь совершенно одинаковый основной рисунок (портрет короля) и отличаться только цветом, ценностью и мелкими деталями в обрамлении. Но может быть и так, что род характеризуется абстрактным понятием, частные случаи которого находят свое воплощение в отдельных видах, например: серии юбилейных марок дома Романовых (отдельные портреты царей), очень многие серии современных советских марок, со временные (вернее, предвоенные) австрийские марки и т. д. Можно найти и закон гомологических рядов: особенно в прежнее время, марки одинаковой значимости (имеющие одинаковое хождение - внутреннее, международное, открытое письмо, бандероль и т. д.) имели одинаковый цвет в разных странах. В филателии понятие рода связано со временем, но и в биологии род вне палеонтологии имеет характер временного разреза пучка линий: к одному роду относятся все марки, объединенные общей идеей и имеющие одновременное хождение. Все сходные роды, преемственно друг друга сменяющие (это особенно характерно, например, для английских и австрийских марок времен Виктории и Франца-Иосифа, где по мере старения монарха рисунок сразу менялся), могут образовать семейство, хотя здесь мы уже отказываемся от признака единовременности, характерного для биологии.
Этот наиболее общий отдел можно наметить пока только в виде краткой программы. Я в своих исканиях исходил из довольно расплывчатых платоновских представлений, навеянных в свое время размышлениями над конкретным эмпирическим материалом и небольшим знакомством с философией (среди биологов особенно важно имя К.К. Шнейдера). Но эти философские идеи должны подвергнуться шлифовке и модификации под влиянием дальнейшего размышления над эмпирическим материалом наряду с углубленным изучением особенно философии Гегеля. Кратко философские проблемы систематики сводятся к следующему (отнюдь не пытаясь охватить все возможные разделы).
А. Проблема реальности таксономических единиц30. Этот вопрос уже был мною затронут в докладе о логических основаниях современных направлений в биологии (печатные тезисы в "Трудах IV Всесоюзного съезда зоологов, анатомов и гистологов" в Киеве в 1930 г.). Я помню, что я там указывал на различные критерии "реальности" таксономических категорий, смотря по природе разобщенности соседних элементов системы, и различал следующие виды разобщенности.
Б. Систематика и морфология. Поскольку основным объектом систематики (хотя и не единственным) является органическая форма, теория и практика систематики связаны теснейшим образом с вопросом о том, что такое органическая форма. На этот вопрос имеется несколько ответов:
Дискуссия по этому вопросу относится уже к морфологии и к проблеме приспособления. Здесь ограничусь лишь тем замечанием, что, несмотря на формальное сходство некоторых выводов химической и биологической систематики (периодическая система элементов; квазипериодические системы многих групп организмов), именно наличие такого формального сходства свидетельствует о наличии глубокого различия реальных сущностей в химии и биологии. Поэтому не удивительно, а, напротив, вполне естественно, что именно механисты и механистические материалисты решительно отрицают возможность рациональной систематики в биологии. В самом деле, организм отличается от химических элементов и соединений двумя признаками: а) чрезвычайной сложностью и, главное, гетерогенностью строения; б) наличием приспособлений. Первый признак не допускает рационализации формы, если отрицать форму как самостоятельную субстанцию в иной плоскости бытия, чем материя (в физическом смысле, не в философском). Второй же признак при придании ему ведущего значения (что обычно у биологов) рассматривает организмы, как находящиеся в состоянии "flexibilitas cerea", восковой гибкости: рационализация формы становится принципиально невозможной. Что иррациональное (ходячее) понимание формы приводит иногда в тупик, об этом свидетельствует, например, откровенное признание д'Арси Томпсона (в его книге "On growth and form"-"Рост и Форма"; Thompson, 1942): возможность превращения очертаний одного вида рыб в другой путем простого преобразования системы координат противоречит эпифеноменальному пониманию формы. Это своего рода доказательство от противного в пользу геометрического характера органических форм. Из антиномии - а) форма - следствие приспособления и б) форма - самостоятельная субстанция, чистая геометрическая форма Скиапарелли - я вижу такой выход: приспособление есть частный случай гармонического строения, которое не противоречит, а вытекает из понятия чистой формы. Для углубления в этот вопрос полезно проштудировать аналогии в языкознании. Язык кажется простым агломератом слов, но этому противоречат такие явления в языкознании:
украинский: хфилософ, хфедор, фист;
русский: философ, Федор, хвост;
русский литературный: цепь, царица, человек, чара;
псковский: чепь, чарича, целовек, цара;
литературный: бегу, бежишь, бежит, бежим, бежите, бегут;
жаргон: бежу, бегешь, бегет, бегим, бегете, бежат;
литературный: хочу, хочешь, хочет, хотите, хотят;
жаргон: хотю, хотит, хочем, хочете, хочут;
В. Метафизика систематики. Всякую метафизику я определяю в ее чисто этимологическом смысле - выхождение за пределы нашего опыта (но отнюдь не имеющую качества принципиально непознаваемого). Здесь мы подходим к самым общим проблемам биологии и философии: вид как идея; организмы - чистые формы; проблема целесообразности как частный случай мировой гармонии, все более теряющей утилитарный характер и все более приобретающей эстетический характер; красота как абсолютная реальность, развитие организмов как воплощение идеи, имеющей конечной целью торжество духа над материей; все это - возрождение подлинного платонизма, главного и (как указал Либманн) единственного серьезного противника дарвинизма, понимаемого как философская система, а не только как эволюционное учение.