© 2000, Annual Reports of the Zoological Institute RAS.
Ludmila A. Kutikova
Zoological Institute, Russian Academy of Sciences, Universitetskaya nab., 1, St. Petersburg, 199034, Russia
Variability and speciation in parthenogenetic organisms is a topical issue of general biological importance. Examples of variability are known in parthenogenetic insects (Suomalainen, 1961), lizards of the genus Chemidophorus (Parker & Selander, 1976), and in angiosperms (Kashin, 1998). Uniqueness of bdelloid rotifers (phylum Rotifera, subclass Archeorotatoria, order Bdelloida; after classification of Markevich, 1990) is that this group of obligatory parthenogenetic worms is very large comprising 360 species belonging to 19 genera and 4 families. Evolutionary pathways of speciation of bdelloid rotifers remain unclear.
Bdelloid rotifers are characterized by apomictic telitoky, i.e. reproduction of only apomictic females. Females have paired gonads and therefore this group of rotifers earlier had the name Digononta. Males have not been found in bdelloids. Doubts concerning obligatory parthenogenesis in bdelloids were expressed by Markevich (1993) and Simakov (1991), who assumed presence of concealed hermaphroditism in rotifers without substantial evidence.
Bisexual reproduction has been only noted in two species of the genus Seison (order Seisonida, subclass Pararotatoria), ectoparasites of marine crustaceans Nebalia. These species are characterized by weakly pronounced sexual dimorphism.
Other more numerous rotifers of the subclasses Hemirotatoria and Eurotatoria (more than 1600 species, 20 families, approximately 100 genera) comprise the group of monogonont rotifers (the former name Monogononta) having one gonad. They reproduce by heterogony, i.e. alternation of sexual reproduction and parthenogenesis, also with participation of apomictic females. Males of these rotifers are dwarf, strongly reduced, but not yet known in all species.
The combination of two types of reproduction and peculiarities of participation of sex process determined the tremendous phenotypical diversity of monogonont rotifers adapted to habitation in different biotopes mostly in inland waters (Fig. 1). Their morphobiological organization is adapted to life in plankton (species of the genera Conochilus, Polyarthra, Asplanchna, Keratella, etc.), in capillary waters of psammon (Dicranophorus, Wierzjeskiella), in periphyton (Collotheca, Floscularia) and even to internal parasitism (Claria, Albertia). The variability of monogononts concerns morphological structures of higher taxonomic levels, and primarily the key structures, i.e. corona and mastax with trophi determining the major evolutionary pathways of the group. They provide for locomotion and feeding of rotifers by interrelated functions. In monogononts key structures are represented by several types of (7 types of corona and 5 types of mastax) reflecting specificity of several types of locomotion and feeding.
Fig.1. Diversity of morphological adaptation in monogononts to different environments. Planktonic: 1 - Synchaeta, 2 - Polyarthra (fam. Synchaetidae); 3 - Asplanchna (fam. Asplanchnidae); 4 - Hexarthra (fam. Hexarthridae); 5 - Conochilus (fam. Conochilidae); 6, 7 - Keratella (fam. Brachionidae). Psammonic: 8 - Dicranophorus, 9 - Wierzejskiella (fam. Dicranophoridae). Phytophylic: 10 - Trichotria, 11 - Macrochaetus (fam. Trichotriidae). Periphytonic: 12 - Collotheca (fam. Collothecidae); 13 - Floscularia (fam.Flosculariidae). Parasitic: 14 - Claria (fam. Clariaidae); 15 - Balatro (fam. Dicranophoridae). From different authors.
Bdelloid rotifers have relatively uniform structure of body adapted to life, in spatially restricted water film of edaphon, in mosses, lichens, leaf and fir litter where they are mostly spread and also in the littoral and profundal of fresh waters, in benthos and among aquatic vegetation. Abruptly changing conditions of the edaphon environment, primarily of humidity and temperature, have led to the development of features of morphological organization similar to those of other soil inhabitants. The helminthoid and homonomous segmentation of the body, dense covers, absence of pigmentation and usually of eyes and also stepping or gliding ways of locomotion are regarded by Ghilarov (1949) as the characteristic features of soil organisms. Bdelloids have false segmentation of the body with a definite number of segments in every section: 3 segments in head and neck, 6 segments in the body, 3-6 segments in foot. Body covers are dense sometimes with cuticular spine-like outgrowths (Dissotrocha). Eyes absent nearly always. Almost all representatives of the families Habrotrochidae, Philodinidae, Philodinavidae, are characterized by stepping locomotion, Adinetidae are characterized by gliding locomotion. Locomotion is performed by well-developed segmented musculature of the body and corona including 3 types: Philodina, Adineta, Abrocha (Melone et al., 1998). In locomotion of bdelloids creeping and swimming modes alternate, therefore their mode of locomotion may be regarded as creeping-swimming (Kutikova, 1970).
Unlike monogononts bdelloids have only one type of mastax - ramate, elements of which are characterized by strong variability (Melone et al., 1998). Apart form the changes in the number of teeth of unci, a considerable variability in the orientation of trophi structures was noted by Melone and Ricchi in the study of jaws by means of electron microscopy.
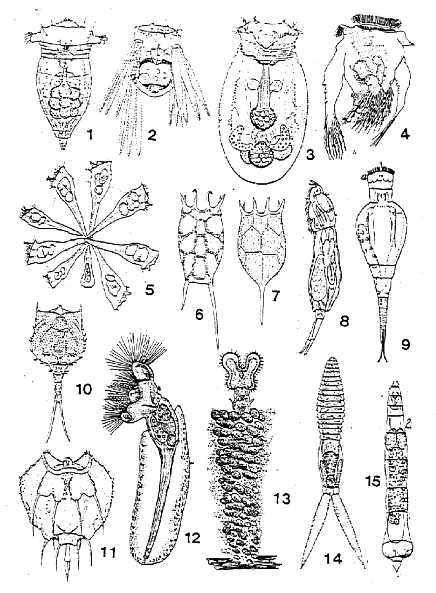
In bdelloids variability observable by means of light microscope is pronounced in the external structures to a larger extent than in the internal structures, e.g. such as the structure of the digestive system: length of oesophagus, width of stomach tube, etc. (Donner, 1965). Only the structure of intestine in Habrotrochidae and other families and also length of tube of intestine in Philodinidae and Philodinavidae belong to characters taxonomically splitting the families. Shape of the body, its covers, structure of the head, parts of corona, foot and its spurs, sometimes of the "house", i.e., the relatively uniform organization of the external structures usually serves as the diagnostic character of the genus. Interspecies characters are even more difficult to define, because they comprise minute details of corona structure and sometimes strongly variable shape of foot spurs. It should be acknowledged that bdelloid rotifers, which can only be studied in vivo, need a more profound anatomo-morphological study using new methods of both microscopy and narcotization. Hetergonetic monogononts in their variability display diversity of morphological organization of the entire body; variability of bdelloids has a different character and concerns characters of the lower taxa (Fig. 2).
Stress situations brought about primarily by the abrupt fluctuations of humidity and temperature of soil environment fostered the development of anhydrobiosis in bdelloids. The questions of ability of bdelloids to fall into anhydrobiosis and to return to active life after hydration, as well as the problem of anhydrobiosis remain unsolved (Crowe, 1971). Anhydrobiosis was probably a stimulus for refusal of bdelloids from sexual reproduction and became biologically analogous to resting fertilized egg.
On the basis of body structure, particularly of corona and mastax hav-ing a more primitive structure as compared to monogononts, Markevich (1993) believes that bdelloids are most similar to the initial ancestral forms of rotifers. The archetype of rotifers theoretically reconstructed by Markevich had muscular type of locomotion and probably was capable of the creeping-swimming mode of locomotion of recent bdelloids. It can be supposed that bdelloids separated from the ancestral form early in their evolution and evolved adapting to capillary and film humid environment of edaphon. This notion contradicts the point of view of Melone and Ricci (1995) who believe that bdelloids evolved from benthic inhabitants of biotopes with unstable environmental conditions.
The branches of bdelloids and monogononts that diverged at an early stage of their development differ in the rates of evolution, the terms of their history being relatively similar. Morphobiological characteristics of bdelloids determined the lower rate of development of the group, and their evolution advanced along the pathway of more particular adaptations not involving characters of higher taxa - subclasses, orders, and families. Intergeneric, interspecific and intraspecific polymorphism in bdelloids is pronounced, but concerns minor external structures.

Fig. 2. Unitypical morphological construction in bdelloids. 1 - Habrotrocha, 2 - Otostephanos, 3 - Ceratotrocha (fam. Habrotrochidae); 4 - Philodina, 5, 6 - Rotaria, 7, 8 - Dissotrocha, 9 - Macrotrachela, 10 - Mniobia (fam.Philodinidae); 10, 11 - Adineta (fam. Adinetidae); 12 - Philidinavus, 13 - Abrochta (fam. Philodinavidae). From different authors.
Therefore it can be assumed that obligatory parthenogenetic bdelloid rotifers invaded terrestrial biotopes on the early stage of their evolution; they are characterized by slow evolution rates as compared to heterogonetic monogononts; the variability of bdelloids is of principally of different character and concerns morphological structures of a lower taxonomic rank.
The studies were carried out with financial support of the Russian Foundation for Basic Research (grant 99-04-49505).
Crowe, J.H. 1971. Anhydrobiosis: an unsolved problem. Am. Nat. 105: 563-573.
Donner, J. 1965. Ordnung Bdelloidea (Rotatoria, Rädertiere). Berlin, Akad. Verl. 297 S.
Ghilarov, M.S. 1949. Osobennosti pochvy kak sredy obitaniya i eye znacheniye v evolyutsii nasekomykh [Characteristic features of soil as a habitat and its significance in the evolution of insects]. Moskva/Leningrad, AN SSSR Publ.. 280 pp. (In Russian).
Kashin, A.S. 1998. Sexual reproduction, agamospermy and species formation in angiosperms. Zh. obshch. Biol. 59 (2): 171-191. (In Russian).
Kutikova, L.A. 1970. Kolovratki fauny SSSR [The rotifer fauna of USSR]. In: Keys of the fauna of USSR. Vol. 104. 744 pp. (In Russian).
Markevich, G.I. 1990. A historic reconstruction of phylogenesis of rotifer as a basis for their macrosystem. In: Kolovratki [Rotifera]: Proceedings of 3rd All-Union rotifer symposium. (L.A. Kutikova. Ed.). pp.140-156. Leningrad, ZIN Publ. (In Russian).
Markevich, G.I. 1993. Evolution of rotifers and the question of their position in the system of Metazoa. Trudy Inst. Biol. vnutrenn. Vod 68 (71): 3-52. (In Russian).
Melone, G. & C. Ricci. 1995. Rotatory apparatus in Bdelloidea. Hydrobiologia 313/314: 91-98.
Melone, G., Ricci, C. & H. Segers. 1998. The trophy of Bdelloidea (Rotifera): a comparative study across the class. Can. J. Zool. 76: 1755-1765.
Parker, E.D. & R.K. Selander. 1976. The organization of genetic diversity in the parthenogenetic lizard Chemidophorus tesselatus. Genetics 84: 791-805.
Simakov, Yu.G. 1991. Osobennosti gametogeneza u kolovratok [Characteristic features of gametogenesis in rotrifers]. In: Voprosy ekologii gidrobiontov [Problems in ecology of hydrocoles]. pp. 133-137. Moskva, VNIIPRKh. Publ. (In Russian).
Juomalainen, E. 1961. On morphological difference and evolution of different polyploid parthenogenetic weevil populations. Hereditas 47: 309-341.