Эмбриональное развитие нервной системы позвоночных |
|||||||||||||||||||||||||||||||||||
Раннее развитиеАвторыТ.А. Цехмистренко |
Нервная система всех позвоночных, включая человека, развивается из элементов наружного зародышевого листка – эктодермы. Этот процесс имеет определенные особенности у представителей разных групп, однако ему свойственны и общие для всех позвоночных закономерности. В период гаструляции у высших позвоночных (у человека это конец 1-й недели развития и совпадает с имплантацией в стенку матки) происходят активные перемещения клеточного материала зародыша. В первой фазе гаструляции образуются два эмбриональных зародышевых листка – эпибласт (верхний листок) и гипобласт (нижний). Клетки эпибласта постепенно расходятся, образуя заполненную жидкостью амниотическую полость. Во второй фазе гаструляции небольшая группа клеток эпибласта, сформировавшая в дне амниотической полости зародышевый щиток, образует первичную полоску и гензеновский узелок. Последующая миграция клеток этих структур вглубь зародыша приводит к формированию среднего листка зародыша – мезодермы. Гаструляция завершается у всех позвоночных образованием трех зародышевых листков: эктодермы, мезодермы и энтодермы, а также формированием осевого комплекса зачатков органов. Особое значение на этом этапе развития принадлежит т.н. головному отростку (нотохорду), формирующемуся из мигрирующих клеток гензеновского узелка. К концу гаструляции формируются и все основные, соответствующие разным группам животных провизорные органы (желточный мешок, амнион, аллантоис, хорион, плацента), выполняющие защитные и питательные функции для эмбриона. Их число в ходе эволюции увеличивается. У человека гаструляция завершается к третьей недели внутриутробного развития. Головной отросток дает начало развитию нотохорды – оси будущего зародыша. Клетки нотохорды и формирующейся затем хорды оказывают индуцирующее влияние на дифференцировку прилежащего к ним участка эктодермы в нервную пластинку и далее в нервную трубку (рис. 1). Как только развивается нотохорд, расположенная над ним эктодерма начинает утолщаться и формирует нервную пластинку, элементы которой интенсивно размножаются и дифференцируются, превращаясь в узкие цилиндрические нейроэпителиальные клетки, отличные от соседних клеток покровного эпителия. Основной причиной формирования нервной пластинки и замыкания ее в нервную трубку является преобразование нейроэпителиальных клеток, связанное с изменением ориентации компонентов их актинового цитоскелета. В результате интенсивного деления и неравномерного роста нейроэпителия происходит его инвагинация с последующим формированием нервной трубки. Эмбриональное развитие ЦНС у млекопитающих и человека обычно разделяют на: эмбриональный (первые 6 недель), фетальный (с 6 до 24 недели) и перинатальный периоды (с 24 недели до рождения) (табл. 1). Таблица 1. Основные стадии развития ЦНС человека (по: ten Donkellar at al., 2006)
|
Содержание
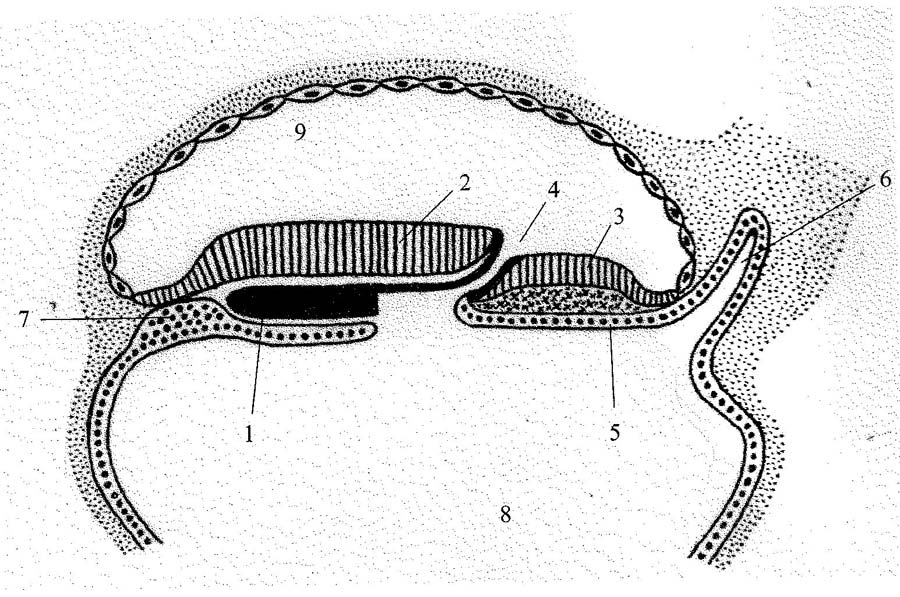
Рис. 1. Схема ранней стадии развития зародыша человека (формирование головного отростка – нотохорды).
1 – головной отросток; 2 – эктодерма, 3 – первичная полоска, 4 – первичная ямка, 5 – энтодерма, 6 – формирующийся аллантоис, 7 – прехордальная пластинка, 8 – полость желточного мешка, 9 – амниотическая полость (по: Данилов, Боровая, 2016, с изменениями) 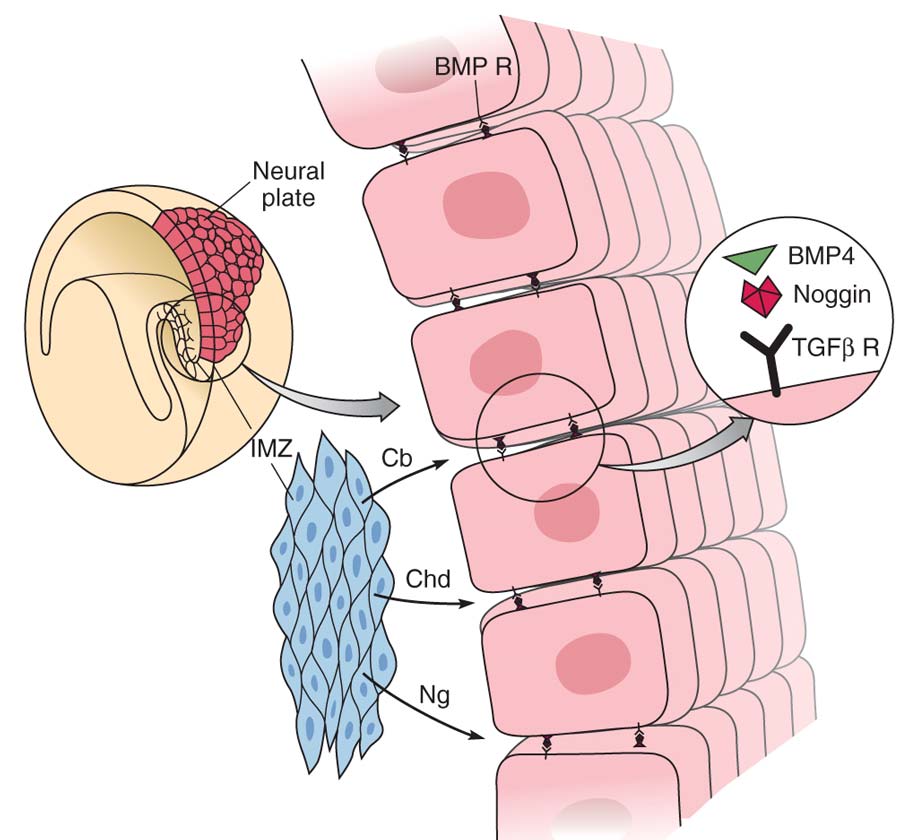
Рис. 3. Первичная индукция формирования нейроэпителия будущей нервной пластинки (по: Development of Nervous System, 2006, с изменениями).
Chd – хордин, Ng – ноггин, IMZ – эмбриональная мезодерма, BPM4 – костный морфогенетический белок, TGFr – рецептор к фактору роста опухолей Рис. 4. Участие ряда сигнальных молекул, транскрипционных факторов и ростовых факторов в формировании нервной пластинки и нервной трубки на ранних этапах эмбриогенеза нервной системы (по: Development of Nervous System, 2006, с изменениями).
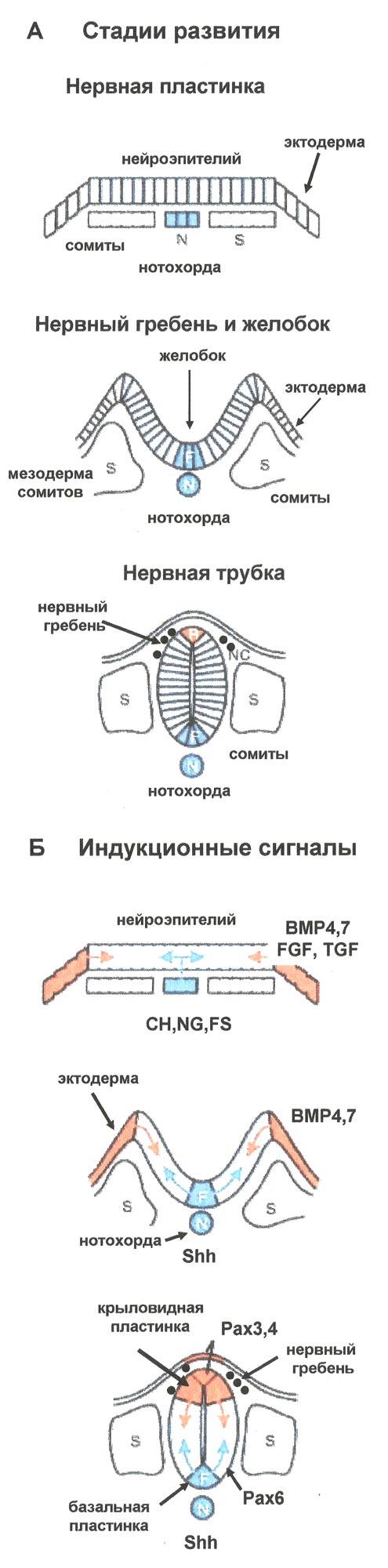
СH – хордин, NG – ноггин, FS – фоллистатин, Shh – sonic hedgehog, BMP – костный морфогенетический белок, FGF – фактор роста фибробластов, TGF – фактор роста опухолей, Pax3, 4, 6 – транскрипционные факторы, S – мезодермальные сомиты, NC – нервный гребень |
|||||||||||||||||||||||||||||||||
Эмбриональный период (1–23 стадии по Carnegie) |
В этот период развития на дорсальной стороне зародыша происходит обособление особого участка нейроэпителия и формирование нервной пластинки (neuronal plate) и начинаются процессы нейруляции. На стадии нейруляции происходит формирование нескольких важных структур нервной системы: образуется нервная пластинка с последующим образованием нервной трубки и нервного гребня (рис. 2). Нейруляция у человека начинается в конце 3-й недели и полностью завершается к концу 4-й недели. Вскоре после образования нервной пластинки (приблизительно на 18-е сутки у человека) она прогибается вдоль продольной оси, ее края приподнимаются и формируются нервный желобок и нервные валики. Позднее края нервных валиков смыкаются по срединной линии и образуется замкнутая нервная трубка. Краниальный и каудальный участки нервной трубки долго остаются незамкнутыми, их называют соответственно передним и задним нейропорами. Передний нейропор закрывается на 23–26-й день развития, а задний – на 26–30-й день. 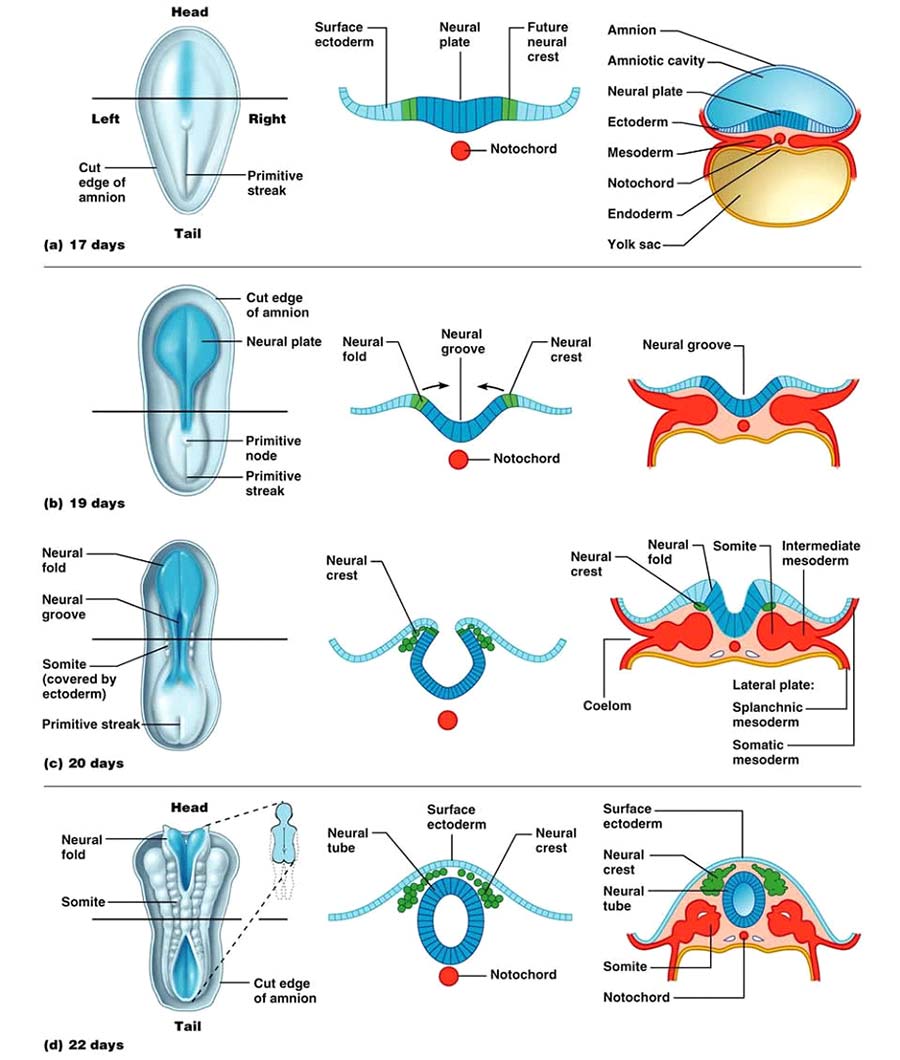
Рис. 2. Ранние стадии развития нервной системы млекопитающих (на примере человека). (по: ten Donkellar at al., 2006, с изменениями).
Процесс нейрональной индукции связан с синтезом ряда биологически активных соединений, которые действуют на формирование нервной пластинки и нервной трубки. На клетки первичной эктодермы действует большое количество сигнальных молекул, индуцирующих процесс образования нейроэпителия и нейрональных стволовых клеток, из которых будут формироваться все элементы нервной ткани. Среди этих факторов необходимо отметить хордин (chordin), ноггин (noggin) и фоллистатин (follistatin), синтезируемые клетками первичной мезодермы, образующей нотохорду (будущая хорда и позвоночник). Они блокируют действие другого морфо-генетического фактора – BMP (bone morphogenetic protein), синтезируемого клетками эктодермы и индуцируют их дифференцировку в направлении образования нейроэпителия нервной пластинки (рис. 3). Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный – крыловидную пластинку, и вентральный – базальную пластинку. Участки мозга, развивающиеся из крыловидной пластинки, содержат ассоциативные и сенсорные ядра, из базальной – моторные и вегетативные. Самая ростральная часть (prosencephalon) не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие производные обеих пластинок – средний, задний, продолговатый – часто объединяют названием «ствол мозга». На этапе формирования нервных желобков дифференцировку вентральной части нервной трубки (базальной пластинки) и развитие мотонейронов оказывает регулирующее влияние фактор Shh (sonic hedgehog) секретируемый сначала нотохордой, а затем хордой и вентральной частью самой нервной трубки. Дорсальную часть нервной трубки (крыловидную пластинку) контролируют морфогенетические белки BMP4 и MBP7, секретируемые клетками эктодермы, и ряд других ростовых и транскрипционных факторов: Pax 3, 4, 6 – транскрипционные факторы, FGF8 – фактор роста фибробластов, GDNF – нейротрофический фактор глии, BDNF, NT3,4 – нейротрофические факторы мозга и др. (рис. 4). Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны:
Вентрикулярная (VZ) зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные или иначе матричные клетки являются по сути нейрональными стволовыми клетками, т.е. предшественниками нейронов и клеток макроглии. Субвентрикулярная зона (SVZ) состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая или мантийная) зона (PZ) состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон – это зона активной миграции и дифференцировки молодых нейронов (нейробластов) и глии (глиобластов). Наружная (маргинальная MZ) зона содержит нервные волокна и отростки находящихся ниже нейронов. Нейробласты достигают мест своего окончательного расположения в структурах ЦНС; утрачивают способность к делению и в дальнейшем дифференцируются в зрелые нейроны. Глиобласты продолжают делиться и дают начало новым популяциям глиальных клеток: астроцитам и олигодендроцитам. |
||||||||||||||||||||||||||||||||||
Образование нервного гребня |
После смыкания валиков и образования нервной трубки, на ее боковых поверхностях выселяется группа клеток, формирующих т. н. нервный гребень (neural crest) (рис. 2). Клетки нервного гребня активно и целенаправленно мигрируют на большие расстояния в зародыше и способны дифференцироваться в разнообразные зрелые элементы тканей и органов. Миграция клеток определяется не только и не столько наличием свободного межклеточного пространства и отсутствием механических преград на пути перемещения, сколько взаимодействием мигрирующих клеток с молекулами межклеточного матрикса (коллаген, ламилин, фибронектин, аминоглюкозгликаны и др.). Формирование фенотипа клеток нервного гребня определяют многочисленные ростовые и дифференцирующие факторы, действующие на клетки гребня по ходу их миграции. В качестве примера можно привести процесс дифференцировки клеток туловищного отдела нервного гребня в нейроны симпатических ганглиев или в клетки хромаффинной ткани мозгового вещества надпочечников (рис. 5). 
Рис. 5. Пути дифференцировки клеток туловищного отдела нервного гребня (по: Gilbert, 2006, с изменениями)
Клетки нервного гребня образуются почти на всём протяжении замыкающейся нервной трубки. Клетки из различных участков нервного гребня дифференцируются не одинаково. Разная дифференцировка клеток наблюдается как по длине гребня, так и по глубине залегания в нем. Из гребня могут образовываться и нервные узлы и большая часть структур черепа. Уникальность этой структуры позволила современным ученым даже считать нервный гребень четвертым зародышевым листком, наряду с эктодермой, энтодермой и мезодермой. Вот неполный список производных клеток нервного гребня:
|
||||||||||||||||||||||||||||||||||
Фетальный период развития (24–46 стадии по Carnegie) |
В этот период развития происходят наиболее значительные события в развитии мозга. Сразу после завершения нейруляции и образования первичных мозговых пузырей, в стенке эмбрионального мозга начинаются интенсивные процессы пролиферации и дифференцировки. Процессы пролиферации и дифференцировки захватывают широкую полосу клеток, расположенных между наружной (базальной) и внутренней (апикальной) поверхностями стенки мозговых пузырей. Они представляют собой нейрональные стволовые клетки (НСК) и развиваются из нейроэпителия нервной пластинки. НСК активно делятся и в процессе прохождения клеточного цикла претерпевают сложные превращения, связанные с последовательными перемещениями в нервной трубке. Перемещение осуществляются путем смешения ядросодержащих отделов клеток внутри формирующихся отростков. Этот процесс получил название интеркинетической ядерной миграции. Ядросодержащие тела клеток двигаются к поверхности нервной трубки, вблизи которой они остаются на некоторое время. Затем ядросодержащие отделы клеток опять перемещаются к вентрикулярной поверхности, после чего НСК втягивают свои отростки и вступают в очередной митотический цикл (M). В результате формируется одно из первых структурных образований развивающейся стенки нервной трубки – вентрикулярный слой (рис. 6а, б). В настоящее время показано, что популяции клеток, составляющих вентрикулярный и формирующийся несколько позднее субвентрикулярный слой, неоднородна. Не все клетки, перемещающиеся в пределах стенки мозга во время митотического цикла, вступают в митоз у вентрикулярной поверхности. В зависимости от присутствия у клеток отростков и характера их контакта с поверхностями стенки мозга выделяют три класса клеток предшественников: монополярные, биполярные и неполярные (рис. 6в). Биполярные клетки (или апикальные предшественники АР) представляют собой либо НСК клетки, либо клетки т. н. радиальной глии (RG), в которые НСК превращаются на самых ранних этапах нейрогенеза. Отличительной особенностью этих клеток является наличие отростков, контактирующих с апикальной и базальной поверхностями стенки мозга на всем протяжении клеточного цикла. Интеркинетические перемещения ядра происходят по этим отросткам и заканчиваются митозом у апикальной поверхности. Монополярные предшественники появляются на более поздних стадиях, когда в стенке мозга формируется субвентрикулярный слой, содержащий также как и вентрикулярный слой НСК. Ядра этих клеток претерпевают интеркинетические перемещения по цитоплазме отростков клеток предшественников, однако в процессе митотического цикла их апикальные или базальные отростки могут терять связь соответственно с апикальной или базальной поверхностями стенки мозга. Митозы происходят как в вентрикулярном слое, так и в верхней области субвентрикулярного слоя. Во внутренних слоях субвентрикулярной зоны у человека недавно были обнаружены клетки предшественники с неполярной морфологией. Характерной чертой этих клеток является ретракция отростков перед митозом и потеря их контакта с апикальной и базальной поверхностью стенки мозга. Они получили наименование «базальные предшественники (ВР). Фактически мы имеем дело с двумя путями образования нейронов в развивающемся мозге. Это – путь прямого нейрогенеза, когда источником нейробластов являются непосредственно НСК и нейрогенная радиальная глия, т. е. апикальные предшественники с моно- или биполярной морфологией, и путь непрямого нейрогенеза, когда источником нейробластов служат промежуточные нейрональные предшественники, являющиеся потомками клеток радиальной глии, т. е. базальные предшественники. Непрямой путь кортикогенеза может выступать в роли быстрого увеличения количества нейронов в условиях ограниченного времени (каждое асимметричное деление радиальной глии через стадию промежуточного нейронального предшественника может давать два – четыре нейрона) и тем самым регулировать площадь и толщину стенки мозга. Таким образом, на первых этапах формирования нервной системы, в стенке эмбрионального мозга формируется широкий слой пролиферирующих нейрональных предшественников разного типа, активность которых в дальнейшем приводит к формированию будущих популяций нервных и глиальных клеток в различных отделах мозга. |

Рис. 6. Классические схемы перемещения клеток в вентрикулярном слое нервной трубки (а, б) и современное представление (в) о гетерогенности нейрональных предшественников. (по: Нейроонтогенез, 1985; Обухов, 2008, Pernavelas et al., 2002, с изменениями)
VZ, ISVZ,OSVZ, MZ – вентрикулярный, внутренний и наружный субвентрикулярный и маргинальный слои стенки мозга; apical, basal – апикальная (внутренняя) и базальная (наружная) поверхности мозговой стенки; Tc, Ts, G1, G2, M – стадии митотического цикла; I – период интерфазы; NB – нейробласт 
Рис. 8. Схема развития мозговых пузырей (A) и формирования головной части зародыша позвоночных (Б) (по: Обухов, Андреева, 2017; Developmental neurobiology, 2005).
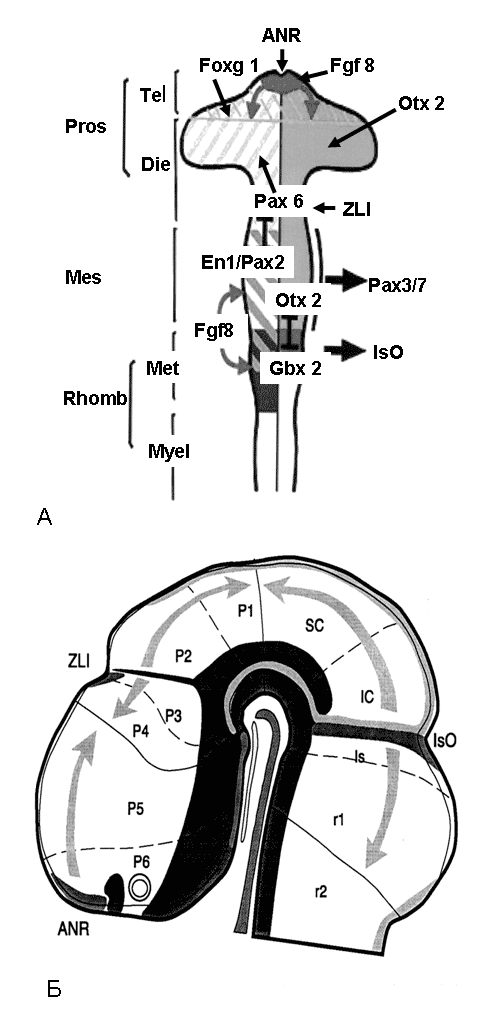
А: сомитомеры и мозговые пузыри костистых рыб и амниот (а), хрящевых рыб и амфибий (б). 1-11 – сомитомеры (будущие туловищные сомиты); 12 – закладка ушной капсулы; 13 – спинальный ганглий; 14, 15 – дорсальный (14) и вентральный (15) корешки спинномозгового нерва; 16 – полость целома; 17 – гипобранхиальная мускулатура; 18 – обонятельный орган, 19 – глазной пузырь; 20, 21 – жаберные щели (20) и дуги (21); III–X, XII – черепно-мозговые нервы Рис. 10. Раннее развитие головного мозга и зоны экспрессии транскрипционных факторов, контролирующих формирование основных отделов головного мозга млекопитающих, вид сверху (А) и сбоку (Б) (Echevarria et al., 2003, с изменениями).
Pros, Tel, Die, Mes, Rhomb, Met, Myel – отделы головного мозга, Р1–Р6 – прозомеры, r1–r2 – ромбомеры, ANR – anterior neural ridge (передний мозговой организатор), ZLI – zona limitans interthalamica organizer (таламический организатор), ISO – isthmic organizer (организатор перешейка), SC, IC – верхние и нижние бугорки четверохолмия, P1–5 – прозомеры, r1–2 – ромбомеры; Pax 3/7, Ent, Otx, Foxg, Gbx – транскрипционные факторы и место их действия; стрелки – направление диффузии регулирующих факторов |
|||||||||||||||||||||||||||||||||
Дифференцировка нервной трубки и формирование основных отделов мозга |
Замыкание нервной трубки начинается в середине зародыша, затем процесс распространяется к головному и хвостовому концам эмбриона, где некоторое время остаются незамкнутыми отверстия – передний и задний нейропоры (рис. 7). 
Рис. 7. Ранние этапы формирования нервной трубки на примере развития мозга человека (по: Nieuwenhuys R. et al., 1999).
A–D – реконструкция вида человеческого зародыша и начальных этапов формирования нервной трубки, E–H – поперечные срезы эмбриона на данных стадиях развития; 1 – эктодерма, 2 – нервная пластинка, 3 – отверстие амниона, 4 – мозговая пластинка, 5 – нервная складка, 6 – нервный желобок, 7 – нервная трубка, 8 – зачаток головного мозга, 9 – передний нейропор, 10 – задний нейропор, 11 – нервный гребень, 12 – крыловидная пластинка, 13 – латеральная пластинка, 14 – базальная пластинка, 15 – полость первичных мозговых желудочков, 16 – зачатки спинальных ганглиев Еще на стадии замыкания нейропоров начинается ростро-каудальная дифференцировка нервной трубки зародыша. Нервная трубка (как полагают, под индуцирующим воздействием хорды) постепенно погружается в мезодерму зародыша и под влиянием мезодермальных сомитов разделяется на сегментарные участки – нейромеры или прозомеры. Сомиты располагаются по сторонам нервной пластинки и вдавливаются в нее, определяя конфигурацию будущих отделов мозга (рис. 8). В дальнейшем головные сомиты сливаются и образуют три основных сегмента: премандибулярный, мандибулярный и гиоидный. Границей головных сегментов служит область ушной капсулы, за которой формируются от 2–3 до 10–12 туловищных сегментов (в зависимости от группы позвоночных). Параллельно формируется система черепно-мозговых нервов. Каждый сегмент иннервируется определенными парами нервов: премандибулярный – терминальным и глазодвигательным нервом (III); мандибулярный – тройничным (V) и блоковым (IV) нервами; гиоидный – отводящим (VI) и лицевым (VII) нервами. Следующие за головными два сегмента иннервируются соответственно языкоглоточным (IX) и блуждающим (X) нервами. Ростральные туловищные сомиты у высших позвоночных иннервируются системой добавочного нерва (XI), включающего в себя разное количество корешков в зависимости от числа туловищных сомитов. Подъязычный нерв (XII), иннервирующий гипобранхиальную мускулатуру, которая развивается из закладки туловищных сегментов, по своей функции аналогичен вентральным (соматомоторным) корешкам спинномозговых нервов, иннервирующих поперечнополосатую мускулатуру туловища и конечностей. Передний конец трубки в конце 3-й недели развития из-за активных процессов пролиферации и миграции нейронов в стенке мозга расширяется и формирует 3 первичные мозговые пузыря. Лежащий краниально пузырь образует первичный передний мозг, Prosencephalon, средний пузырь – первичный средний мозг Mesencephalon, а из третьего пузыря развивается первичный задний мозг Rhombencephalon. Далее располагаются структуры формирующегося спинного мозга – Medulla spinalis (рис. 9).  Рис. 9. Развитие мозга человека (по: Шаде, Форд, 1976).
А–Б – стадии трех (а) и пяти (б–д) мозговых пузырей; вид сверху (А) и сбоку (Б); 1–3 – первичные: передний (1), средний (2), ромбовидный (№) мозг; 4 – закладка спинного мозга, 5 – глазной бокал, 6–10 – отделы мозга: конечный (6), промежуточный (7), средний (8), задний (9), продолговатый (10); 11 – полушария конечного мозга, 20 – мозжечок, 22 – спинной мозг, V–IX – черепно-мозговые нервы. Стрелки – изгибы нервной трубки (с. и. – среднемозговой, ш. и. – шейный, м. и. – мостовой) Спинной мозг образуется из каудальных отделов нервной трубки. Он представляет собой часть ЦНС, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегментарность. После формирования мозговых пузырей в нервной системе начинаются сложные процессы внутренней дифференцировки и роста. Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный – крыловидную пластинку, и вентральный – базальную пластинку. Участки мозга, развивающиеся из крыловидной пластинки, содержат сенсорные ядра, из базальной – моторные и вегетативные. Ростральная часть нервной трубки не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие производные обеих пластинок – средний, задний, продолговатый – часто объединяют названием «ствол мозга». Изменения в развитии нервной трубки сопровождаются образованием нескольких изгибов на границах закладки различных отделов мозга. В течение первых двух месяцев эмбрионального развития образуется основной (среднемозговой) изгиб, когда передний и промежуточный мозг загибаются вперед и вниз. Затем формируется еще два (шейный и мостовой) изгиба. Одновременно первый и третий первичные мозговые пузыри разделяются каждый на два. Наступает стадия пяти мозговых пузырей. Самым ростральным становиться конечный мозг (Telencephalon), затем – промежуточный (Diencephalon). За промежуточным идет средний мозг (Mesencephalon). Первичный задний мозговой пузырь разделяется на задний мозг (Metencephalon) и продолговатый мозг (Medulla oblongata). Прозенцефалон включает производные первых шести прозомеров (нейромеров) P1–P6. Из структур Р1 в дальнейшем формируется средний мозг. Прозомеры Р2 и Р3 развиваются соответственно в таламус и преталамус. Из прозомеров Р4–Р6 развивается конечный мозг и гипоталамус. Из более каудальных сегментов нервной трубки (ромбомеров) развиваются структуры ствола и спинного мозга. После формирования мозговых пузырей (5–10 недели развития) в структурах формирующейся нервной системы происходят сложные процессы внутренней дифференцировки и роста различных отделов головного и спинного мозга. Формирование отделов мозга находится под контролем т. н. «вторичных организаторов» – групп клеток, синтезирующих ряд морфогенетических факторов, градиент концентрации которых определяет направление миграции и дифференцировки разных структур мозга (табл. 2; рис. 9, 10). Таблица 2. Гены и продукты их экспрессии, контролирующие различные процессы развития
отделов и структур головного мозга (по: Обухов, 2008).
Развитие зачатка переднего мозга контролирует небольшая группа клеток, расположенная на верхушке нервной трубки и названная передним мозговым организатором (ANR – anterior neural ridge) и клетки на границе второго мозгового пузыря – zona limitans interthalamica (ZLI). Структуры среднего, заднего, продолговатого мозга и верхние сегменты спинного мозга контролируются еще одним организатором – isthmic organizer (ISO). В перинатальный период заканчивается формирование внутренней структуры мозга. Начинается активная миелинизация головного и спинного мозга. Однако эти процессы не заканчиваются с рождением. Показано, что достаточно долго (месяцы и годы) после рождения происходит созревание и дифференцировка нервных структур и проводящих трактов. Более того, в настоящий период стало ясно, что во взрослый период происходит образование новых популяций нейронов и глиальных клеток за счет сохранения в мозге популяций НСК в структурах головного мозга. |
||||||||||||||||||||||||||||||||||
Список литературы |
|
||||||||||||||||||||||||||||||||||