Краткий обзор организации и эволюции
|
||
АвторРедактор |
Становление типа хордовых (Chordata) шло в докембрии в период, когда примитивные оболочники (подтип Tunicata) переходили к неотении и потере прикрепленной стадии. Примитивная нервная система трубчатого типа сформировалась у предков хордовых довольно рано и сразу как единое целое. У асцидий – наиболее характерных представителей подтипа оболочников – на соответствующих стадиях онтогенеза также развивается нервная трубка, расположенная в длинном хвостовом отделе личинок над хордой и кишечником. Передний конец нервной трубки увеличен и образует так называемый мозговой пузырёк, с которым связаны примитивные рецепторные органы – многоклеточный светочувствительный глазок и одноклеточный статоцист – орган гравитации. На более поздних стадиях онтогенеза асцидий происходит регрессивный метаморфоз, связанный с переходом взрослых животных к «сидячему» образу жизни на морском дне. В процессе метаморфоза основная часть нервной трубки атрофируется и у взрослого животного сохраняется лишь надглоточный (мозговой) ганглий, возникающий из спинной стенки мозгового пузырька личинки. |
Содержание
|
Появление трубчатой
|
У всех хордовых зачаток нервной системы располагается на спинной стороне зародыша, образуя нервную пластинку над зачатком хорды. Появление трубчатой нервной системы у первичных бесчерепных (подтип Acrania) было связано с развитием осевой мускулатуры – т.е. «миохордального комплекса», иннервация которого осуществлялась за счет интраэпителиальных нервных сплетений. Эти первичночувствующие интраэпителиальные клетки, обособляясь от покровного эпителия, погружались в стенку туловища, формируя первичную нервную пластинку. Впоследствии боковые края нервной пластинки утолщаются и приподнимаются в виде пары нервных (или медуллярных) валиков, так что средняя часть нервной пластинки оказывается вогнутой. Так формируется нервный желобок, тянущийся вдоль спинной стороны зародыша. Дальнейшее разрастание нервных валиков кверху и к средней линии завершается их смыканием друг с другом, при этом желобок превращается в нервную трубку. Клетки покровной эктодермы, примыкающие сбоку к нервному желобку, интенсивно размножаются в направлении к средней линии, а эктодерма смыкается над нервной трубкой. Покровное, эктодермальное происхождение нервной трубки подтверждается также тем, что довольно долго на ростральном и каудальном полюсах зародыша позвоночных сохраняется отверстие, соединяющее полость нервной трубки с наружной средой – нейропоры. Трубчатый характер нервной системы оказался настолько эффективным, что по существу не подвергался принципиальным изменениям за всю историю эволюции хордовых. |
|
Появление
|
Еще в 1868 году Ф.В. Овсянников доказал, что уже на этапе бесчерепных (Cephalochordata), представленных ныне ланцетниками, начинается дифференциация нервной трубки на головной и туловищный отделы. Это было связано с концентрацией на переднем конце туловища древних бесчерепных первичных рецепторных образований и их центров. В ходе дальнейшего эволюционного развития бесчерепные были вытеснены первичными черепными (Craniata), имевших уже достаточно развитые органы чувств и дифференцированный головной и спинной мозг. Позвоночные животные разделяются на две крупные ветви: бесчелюстные (Agnatha), которые представлены сейчас одним классом Круглоротые (Cyclostomata) и челюстноротыми (Gnathostomata), куда входят все остальные позвоночные. Исходной группой для обеих ветвей были телодонты (подкласс Thelodonti класса Pteraspidomorphi) – некрупные беспанцирные формы, обитавшие в прибрежных участках морей и в лиманах. Примитивные бесчелюстные были придонными, проходными обитателями дельтовых водостоков и лиманов. Головной мозг первичных бесчелюстных был вытянут в одной плоскости, обонятельные лопасти не были обособлены от переднего мозга, мозжечок и промежуточный мозг были асимметричны и уже достаточно хорошо развиты. Задний мозг был связан с октаво-латеральной сенсорной системой, средний – со зрительной, а передний – с обонятельной. Таким образом, уже на этой стадии эволюции головной мозг дифференцировался на ряд отделов, возникновение и развитие которых, как полагают, теснейшим образом связано с развитием важных для ориентации в пространстве дистантных сенсорных систем, к которым главным образом относят зрительную и обонятельную системы. |
|
Круглоротые |
Круглоротые (Cyclostomata) произошли от нектонных бесщитковых бесчелюстных подкласса Anaspida, некоторые формы которых, имевшие лишь кератиновые чешуи, приспосабливались к использованию тел погибших панцирных бесчелюстных как временных убежищ и источников пищи. Сформировавшиеся таким образом миноги (подкласс Petromyzones) переходили к эктопаразитизму и полухищничеству в пресных и солоноватых водоемах, а миксины (подкласс Myxini) — к особой форме хищничества в море. Разделение двух ветвей круглоротых произошло еще до начала карбона. На этом длительном пути эволюции от Anapsida к типичным миногам и, особенно, к миксинам могло произойти некоторое вторичное упрощение в строении и функционировании их центральной нервной системы. Тем не менее, круглоротые являются первым классом позвоночных, у которых сформированы пять основных отделов головного мозга позвоночных, включая человека (рис. 1). |
|
Отделы мозга
|
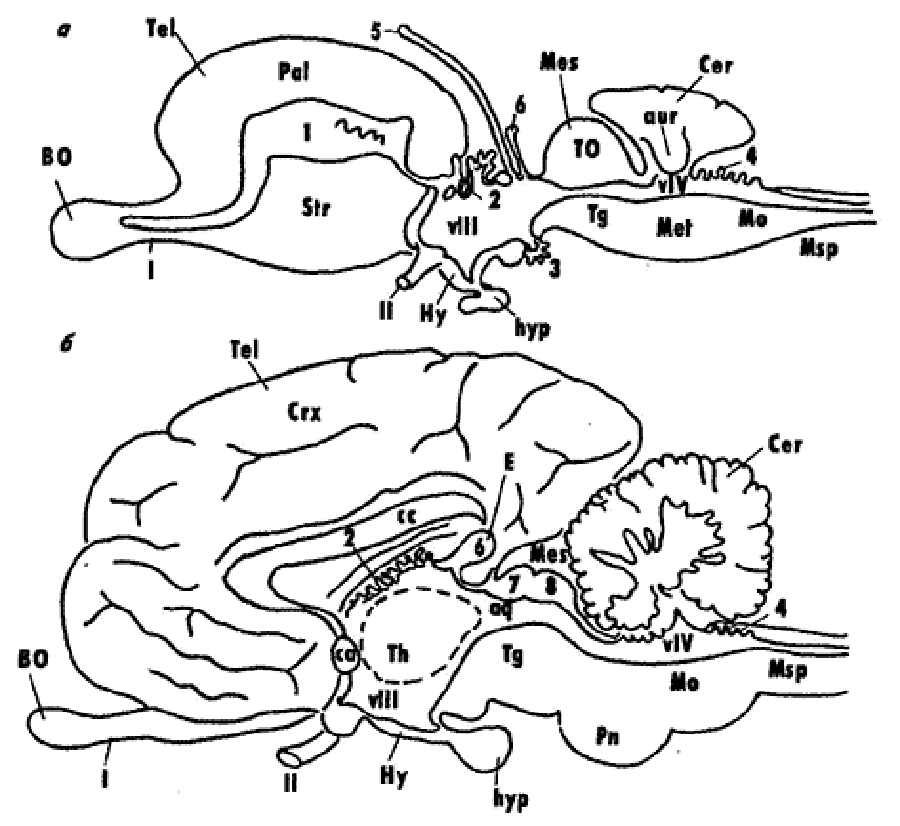
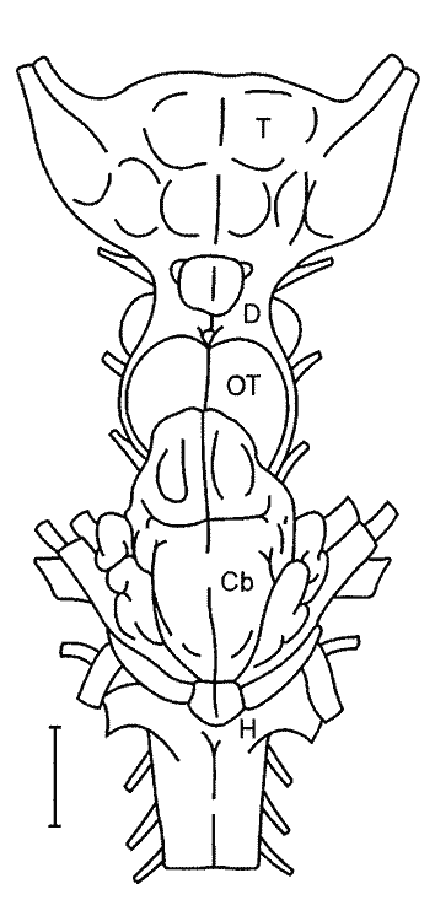
Самым ростральным отделом нервной трубки становится конечный мозг — Telencephalon. Каудальнее располагается промежуточный мозг — Diencephalon, за которым следует средний мозг — Mesencephalon. Первичный задний мозг разделяется на вторичный задний мозг — Metencephalon и продолговатый — Medulla oblongata, переходящий в спинной мозг — Medulla spinalis (рис. 2).
Рис. 2. Схема общей организации головного мозга низших и высших позвоночных (по: Андреева, Обухов, 1999): сосудистые сплетения конечного (1), промежуточного (2 и 3), заднего (4) мозга; 5 – теменной орган, 6 – пинеальный орган (эпифиз); верхние (7) и нижние (8) бугорки четверохолмия среднего мозга; VIII и VIV – желудочки промежуточного и заднего мозга; обонятельный (I) и зрительный (II) черепно-мозговые нервы; BO – обонятельная луковица, Tel – конечный мозг, Crx – кора полушарий, cc – мозолистое тело, Str – стриатум (базальные ядра), Hy – гипоталамус, hyp – гипофиз, Th – таламус, E – эпифиз, Mes – средний мозг, Tg – тегментум, Pn – мост, Mo – продолговатый мозг, TO – тектум оптикум среднего мозга, Msp – спинной мозг, Pal – корковые формации конечного мозга низших позвоночных (паллиум).
Спинной мозг - Medulla spinalis образуется из каудальных отделов нервной трубки. Он представляет собой часть центральной нервной системы (ЦНС), в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных - трубчатый характер строения и сегментарность. Продолговатый мозг - Medulla oblongata является наиболее каудальным отделом головного мозга, плавно переходящим в спинной. Условной границей головного и спинного мозга считают место выхода первой пары спинномозговых нервов. Наиболее значительное сходство со спинным мозгом наблюдается в каудальных отделах продолговатого мозга и выражается в расположении серого и белого вещества, а также в центральном положении мозгового канала. Последний в ростральном направлении расширяется и превращается в ромбовидную или треугольную полость – IV мозговой желудочек, ventriculus quartus. Крыша его истончена и составлена лишь эпендимным эпителием, снаружи от которого у высших позвоночных лежит сильно васкуляризованная мягкая оболочка мозга. У низших позвоночных серое вещество представляет собой тяжи, или столбы, нервных клеток, идущие вдоль продолговатого мозга, тогда как у высших позвоночных намечается четкая тенденция к разделению этих тяжей на отдельные ядра. Задний мозг - Metencephalon у всех позвоночных представлен мозжечком - Cerebellum, форма, размеры и соотношение частей которого сильно варьируют у представителей разных групп позвоночных и в целом коррелируют со степенью совершенства их моторных реакций. В вентральной и боковых стенках заднего мозга, слабо развитых у низших позвоночных, у млекопитающих формируется мощная система проводящих трактов и ядер, образующая самостоятельный отдел — мост — Pons. Полостью заднего мозга являются ростральные отделы IV мозгового желудочка. Средний мозг — Mesencephalon. В составе среднего мозга выделяют производное крыловидной пластинки — крышу среднего мозга (tectum mesencephali) и производное базальной пластинки — покрышку (tegmentum mеsencephali). У высших позвоночных вследствие появления новых отделов мозга и их связей на месте зрительных долей развиваются структуры бугорков четверохолмия — colliculi superiores et inferiores и возникают дополнительно ножки мозга (pedunculi cerebri), представляющие собой систему трактов, связывающих вышележащие отделы ЦНС с нижележащими. Полостью среднего мозга являются остатки полости среднего мозгового пузыря - водопровод мозга (aqueductus cerebri). Промежуточный мозг - Diencephalon включает в себя лежащий дорсально эпиталамус (epithalamus), занимающий срединную часть таламус (thalamus) и расположенный вентрально гипоталамус (hypothalamus). У высших позвоночных выделяют еще субталамус (subthalamus). Последний включает крупные нервные центры — латеральные и медиальные колечатые тела - corpus geniculatum laterale et medialis. Следует отметить, что таламус промежуточного мозга претерпевает наибольшие структурные преобразования в ряду позвоночных, что во многом обусловлено развитием связей это отдела мозга с прогрессивно развивающимся конечным мозгом. Конечный мозг — Telencephalon представляет собой наиболее сложный отдел головного мозга. Формирование его в эволюции различных групп позвоночных шло двумя принципиально разными путями (см. ниже). Максимального развития полушария конечного мозга достигают у высших млекопитающих – приматов и человека. В конечном мозге выделяют лежащие дорсально паллиальные Pallium и расположенные вентрально субпаллиальные Subpallium отделы. Первые представлены кортикальными формациями, которые разделяются на древнюю Paleocortex, старую Archicortex и новую Neocortex кору. Подкорковые образования конечного мозга так же, как и кора, проходят у позвоночных животных сложный путь эволюции и имеют в своем составе филогенетически разные отделы - палеостриатум, архистриатум и неостриатум. |
|
Теория
|
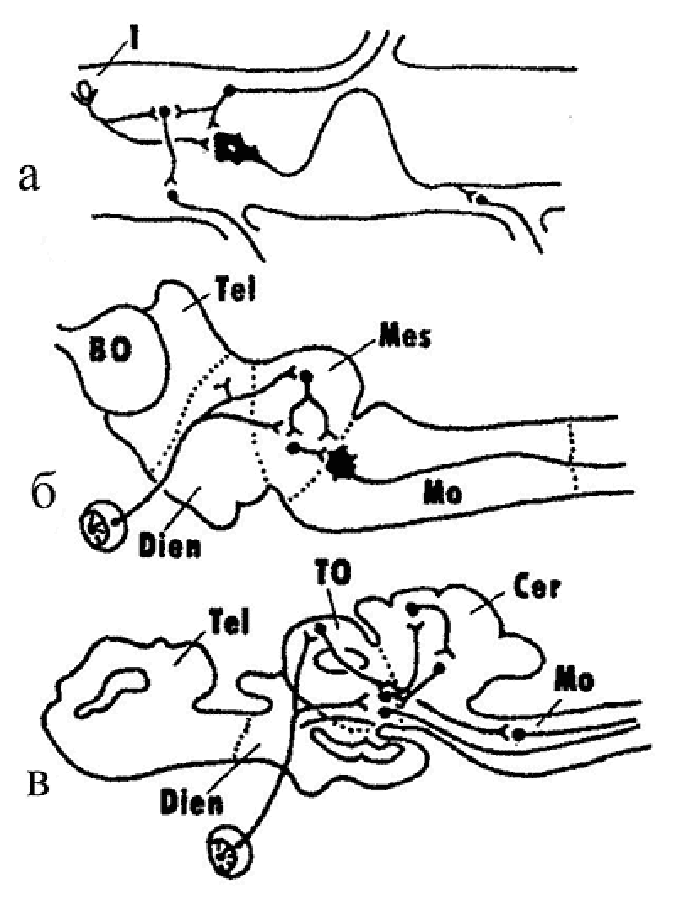
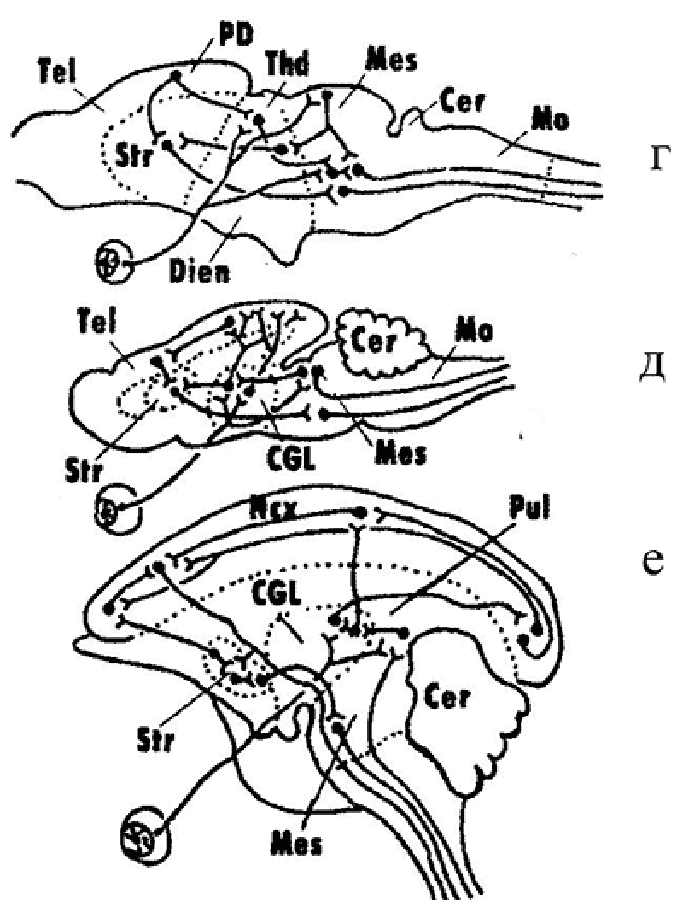
В 50–70-е годы чл.-корр. АН СССР А. И. Карамян – крупный советский нейрофизиолог-эволюционист, ученик и последователь идей Л. А. Орбели сформулировал теорию критических периодов в филогенезе ЦНС позвоночных. В основу теории легли данные о том, что развитие головного мозга позвоночных происходило в эволюции от диффузных, неспециализированных форм к специализированным формам структурной и функциональной организации. При этом в определенные – «критические» (по определению А. И. Карамяна) – периоды развития позвоночных устанавливается строгая корреляция между степенью специализации структур ЦНС и уровнем условно-рефлекторной деятельности. Этот процесс осуществляется путем «надстройки» филогенетически молодых, прогрессивных отделов ЦНС над «старыми» отделами мозга, которые при этом теряют часть свойственных им ранее функций и систем связей (рис. 3). Исследование структурно-функциональной организации нервной системы ланцентика и круглоротых позволило отнести эту группу позвоночных к первому критическому этапу эволюционного развития ЦНС позвоночных (рис. 3а, б). Как было отмечено, у ланцетника признаки цефализации почти отсутствуют. Мозг имеет диффузную организацию, головной мозг как отдел ЦНС еще не сформирован. Этот уровень интеграции получил наименование спинального. У круглоротых (миног), в отличие от бесчерпных, морфологически уже выражены все основные отделы головного мозга, однако элементы диффузной организации мозга еще сохраняются. Преимущественное развитие у них продолговатого и среднего мозга обусловило наименование этого этапа интеграции как бульбо-мезенцефального. Об остальных критических этапах развития ЦНС позвоночных будет рассказано дальше при описании ЦНС соответствующих групп животных. |
|
Первые
|
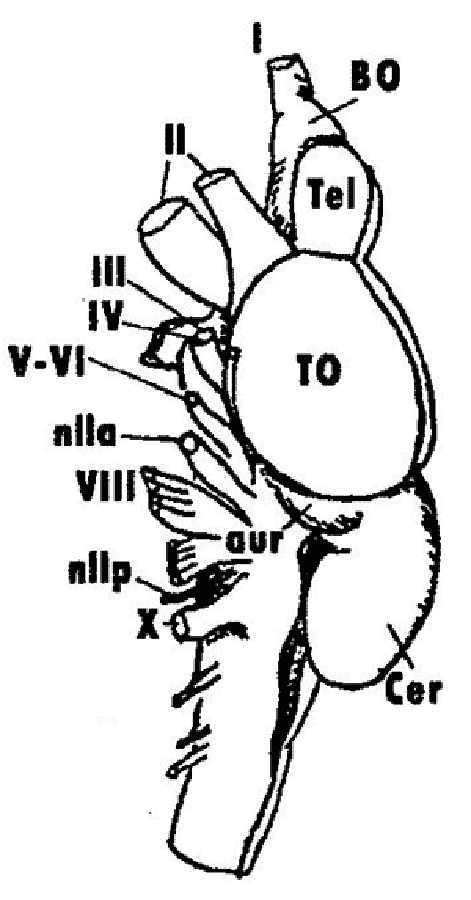
Преобразование первых жаберных дуг в челюстной аппарат, служащий для активного схватывания пищи, и развитие парных плавников дали первым челюстноротым значительные эволюционные преимущества перед бесчелюстными. Способность к активному питанию резко повысила их жизнедеятельность, позволила завоевать водную среду и дала начало одному из самых многочисленных и разнообразных по жизненным формам надклассу Рыб (Pisces). Наиболее древним и ныне вымершим классом рыб являются Панцирные рыбы (Placodermi). В начале девонского периода появились Хрящевые рыбы (класс Chondrichthyes). Они в большей степени, чем панцирные, сохранили облик и основные особенности организации предковых форм. Пластиножаберные (подкласс Elasmobranchii) являются крупнейшей группой хрящевых рыб, которые в палеозое были весьма многочисленны в солоноватых водах морей и лиманов, а в кайнозое активно заселили моря и океаны. Они разделяются на акул и скатов, представляющих собой разные ступени эволюции рыб. Эласмобранхи со скатообразным телом не раз возникали от акулоподобных предков. В настоящее время консервативными реликтами палеозоя являются плащеносные акулы, с юры известны морские ангелы и гребнезубые акулы, с мела – пилоносы, пилы-рыбы, разнозубые акулы. Особую, в целом прогрессивную, группу пластиножаберных рыб составляют ламноидные акулы, кархарины и некоторые ветви скатов. Они играют значительную роль в экосистемах морей, занимая вершины трофических пирамид. Современные Хрящевые рыбы – процветающая и ныне группа рыб, заселившая обширные районы Мирового океана и некоторые пресноводные водоемы и насчитывающая в своем составе около 50 семейств и несколько сот видов акул, скатов и химер. Головной мозг современных акул и скатов отличает, в первую очередь, четкое разделение всех отделов и значительное развитие конечного мозга (рис. 4). Отличное обоняние, зрение, совершенная двигательная система коррелируют с развитием соответствующих отделов головного мозга (обонятельных долей, конечного, среднего мозга, мозжечка). Индекс телэнцефализации (отношение веса конечного мозга к весу всего мозга) достигает у ряда акул значений, сходных с высшими позвоночными. |
|
Костные рыбы
|
Костные рыбы появились в девонском периоде палеозойской эры независимо от хрящевых рыб. Предки костных рыб очень рано разделились на подклассы лучеперых рыб (Actinopterygii), многоперов (Polypterigii) и мясистолопастных (Sarcopterygii). Древние лучеперые рыбы из надотряда палеонисков (Palaeonisci) уже в палеозое очень широко дивергировали. Так, от одного из стволов палеонисков серией ветвей возникли цельнокостные рыбы - надотряд Holostei, обитавшие в солоноватых и пресных водах мезозоя. Ныне эта группа представлена лишь двумя семействами консервативных реликтов мелового периода – панцирными щуками и ильными рыбами. Уникальность группы Лучеперых рыб состоит в том, что у них при сохранении общего для позвоночных животных плана строения головного мозга сформировался особый эвертированный тип строения конечного мозга. При этом типе строения в полушариях конечного мозга отсутствуют латеральные мозговые желудочки, характерные для инвертированного типа конечного мозга всех остальных позвоночных от круглоротых до млекопитающих (рис. 5). 
Рис. 5. Два типа развития конечного мозга позвоночных (по: Андреева, Обухов, 1999): а – ранняя стадия развития полушарий; б - инвертированный и в - эвертированный типы строения конечного мозга; 1 – стриатум, 2 – септум; 3-5 – соответственно латральная, дорсальная и медиальная зоны паллиума (коры полушарий).
За счет параллельных преобразований не менее чем трех ветвей древних цельнокостных рыб постепенно сформировались ныне живущие костистые рыбы (надотряд Teleostei) (рис. 6). Появившись в мелких лиманных водах палеозоя, они постепенно замещали цельнокостных рыб, а к концу раннего мела широко распространились в морях и пресных водах Считается, что Хрящевые ганоиды (надотряд Chondrostei) возникли от особого ствола палеонисков и развивались по пути фетализации - задержки ряда признаков ранних стадий онтогенеза до его более поздних стадий. Эта группа рыб уже в середине мела разделились на ветви осетровых рыб и веслоносов (рис. 7). По мнению А.И. Карамяна с значительным развитием у рыб среднего мозга и мозжечка связан второй критический этап в развитии ЦНС позвоночных. Именно в мозжечке рыб концентрируются проекции всех сенсорных систем (соматосенсорной, органов боковой линии, вестибулярной, зрительной и слуховой). Ведущим центром интеграции в ЦНС у рыб становится средний мозг и мозжечок. Это – мезенцефало-церебеллярный тип интеграции (рис. 3в). Структуры переднего мозга в морфо-функциональном отношении у большинства представителей рыб еще не развиты и выполняют, в основном, функцию обонятельного центра. Хотя, по современным данным, хрящевые рыбы не совсем попадают под характеристику этого этапа развития нервной системы. У многих акул высшим центром интеграции становиться конечный мозг, сближая их с представителями высших классов позвоночных. |
|
Происхождение
|
Исходной группой костных рыб, от которых произошли первые наземные позвоночные (Tetrapoda) являются древние кистеперые рыбы из группы рипидистий (Rhipidistia). Как сейчас признается палеонтологами, климатические условия в девонском периоде (времени выхода позвоночных на сушу) были очень изменчивы. Это способствовало значительной адаптивной радиации саркоптеригий. Часть рипидистий активно осваивала пресноводные водоемы, другие переходили к жизни в морских лагунах, которые могли опреснятся и периодически высыхать. Все это могло привести к необходимости приспосабливаться к полуводному образу жизни и к приобретению новых свойства, ставших в дальнейшем предпосылками выхода на сушу предков амфибий. Следует подчеркнуть, что формирование многих признаков, характерных для амфибий, происходило у их предков еще в водной или полуводной среде. Они оказались способными относительно быстро занимать случайно освобождающиеся места в экосистемах водоемов, замещая кистеперых в их исходной водной и, отчасти, полуводной среде. Таким образом, приспособление к крайне необычным для рыб условиям жизни привело в эволюции позвоночных к выработке адаптаций, обеспечивших освоение новой для них наземной среды. Приобретение этих адаптаций происходило среди филетически различных групп рипидистий, и ряд «полуамфибий» так и остались слепыми ветвями эволюции. Строгой и единственной линии, ведущей от рыб к амфибиям, не существовало. |
|
Земноводные
|
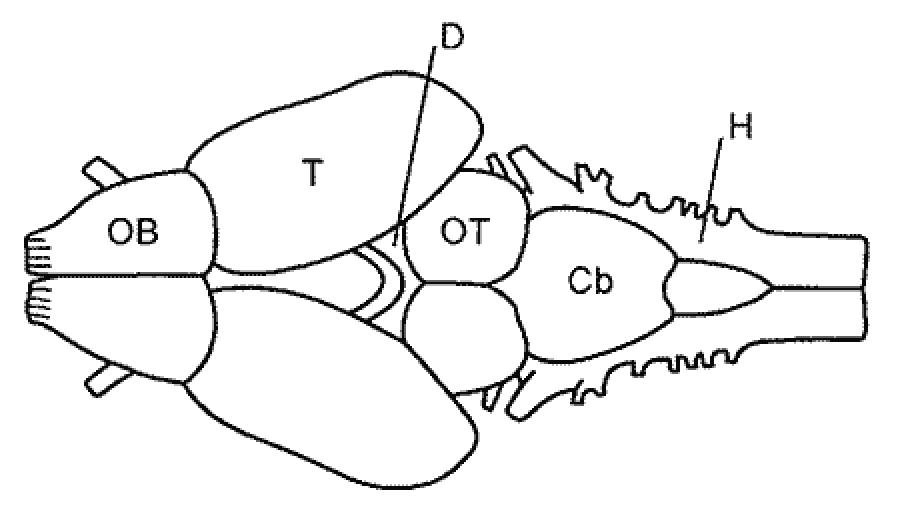
Современные амфибии, составляющие надотряд Lissamphibia, представлены тремя отрядами: хвостатыми (Urodela), бесхвостыми (Anura) и безногими (Apoda). За этими тремя группами признается монофилетическое происхождение, причем бесхвостые амфибии являются потомками ствола, более 350 млн. лет развивающегося обособленно от хвостатых и безногих. Их формирование происходило в палеозое от достаточно сильно различающихся предковых групп, обитавших на разных массивах суши, относящихся в настоящее время, соответственно к Северному и Южному полушарию. Переход к наземному образу жизни способствовал значительной дифференцировке и усложнению как органов чувств, так и связанных с ними отделов ЦНС. Начиная с земноводных, в эволюции ЦНС позвоночных происходят коренные изменения в степени развития и соподчинении отделов головного мозга (рис. 8). Именно с амфибиями, а в дальнейшем и с рептилиями связан третий критический этап в развитии ЦНС позвоночных. У амфибий в связи с переходом к новым условиям жизни на суше произошли существенные изменения в морфо-функциональной организации организма и ЦНС по сравнению с костными рыбами. Редуцируется целый ряд структур, связанных с водным образом жизни (например, органы боковой линии); развиваются новые, филогенетически молодые системы связей (слуховые органы и центры); более четко дифференцируются отделы таламуса; в полушариях закладываются формации неокортекса; устанавливаются прямые таламо-кортикальные связи. Вся система мозговых центров главных сенсорных систем начинает перемещаться из ствола, мозжечка и среднего мозга в верхние отделы - промежуточный и конечный мозг, формируя диенцефало-теленцефальный уровень интеграции (рис. 3г). Главным направлением цефализации позвоночных становиться всемерное развитие конечного мозга (теленцефализация), достигающая высшего развития у млекопитающих и человека. |
|
Высшие
|
Эволюционная ступень высших позвоночных амниот (Amniota) включает две ветви, одна из которых, зауропсидная, содержит классы Рептилии (Reptilia) и Птицы (Aves), а другая, терапсидная, включает классы Зверообразные (Theromorpha) и Млекопитающие (Mammalia). Данные палеонтологии, сравнительной морфологии и физиологии показывают, что обе эти ветви могли достичь уровня амниот вполне самостоятельно, при этом ряд важных признаков организации сформировался еще на «амфибийной» стадии развития. Усложнение и интенсификация функций органов чувств и органов движения у амниот, изменения в их поведении в новой более разнообразной среде обитания (амниоты полностью освоили наземную, водную и воздушную среду) без сомнения связаны с развитием у них более высоко организованной ЦНС. |
|
Рептилии
|
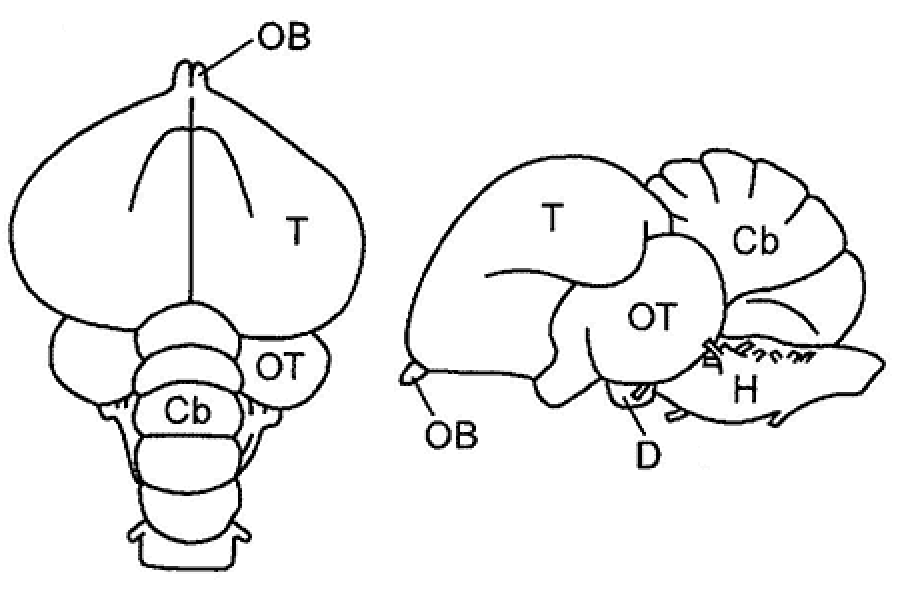
Среди современных рептилий относительно прямыми потомками первых рептилий котилозавров (подкласс Cotylosauria) могут рассматриваться черепахи (подкласс Testudines), стоящие во многом особняком от остальных рептилий (рис. 9). От мелких, подвижных насекомоядных котилозавров (хищных по способу питания) ведет свое начало другая группа рептилий – чешуйчатые (подкласс Lepidosauria). Сохранившаяся c начала мезозоя и до наших времен реликтовая форма из отряда клювоголовых (отряд Rhynchocephalia) – гаттерия обитает в довольно простой биотической среде. Широкая адаптивная радиация ящериц (отряд Lacertilia) современной фауны происходила в основном с середины мезозоя. Нынешние группы гекконовых, агамовых, сцинковых, настоящих ящериц, варановых и других характеризуют разные и, подчас, параллельные ветви развития рептилий. Змеи (отряд Serpentes) являющиеся филогенетически самым молодым отрядом рептилий, прошли в меловом периоде стадию роющих предков, имевших конечности. На вторичность процесса утраты конечностей у змей указывает, в частности, тот факт, что у них сохраняются в рудиментарном виде вся мышечная система конечностей и соответствующие нейронные объединения спинного мозга. В группе безногих ящериц (отряд Amphisbaenia) утрата конечностей и переход к роющему образу жизни в более простой среде обитания также вторичен по отношению к их активным и подвижным ящеричным предкам. Архозавры (подкласс Archosauria) или высшие рептилии также возникли непосредственно от котилозавровых предков еще на «прорептилийной» стадии развития. Уже в мезозое члены этой крупной группы рептилий (типичными представителями которой были динозавры) обладали сложным поведением, некоторые из них приобрели определенный уровень теплокровности (отряд летающих ящеров Pterosauria). Единственные из оставшихся в настоящее время архозавров – крокодилы отряд Crocodilia) являются лишь умеренно высокими по уровню организации в группе архозавров. Наибольшим числом примитивных черт организации среди современных крокодилов обладают аллигаторы и кайманы. С позиций эволюционной нейроморфологии рептилии представляют исключительный интерес, поскольку именно в этом классе позвоночных закладываются те признаки организации ЦНС, которые дали начало развитию высших центров головного мозга птиц и млекопитающих. В настоящее время на смену гипотезе происхождения птиц (класс Aves) от ранних архозавров (отряд Thecodontia) пришла гипотеза их возникновения от группы мелких хищных динозавров из отряда Saurischia. Последние обладали бипедией (передвигались на двух нижних (задних) конечностях), имели относительно большой объем головного мозга и ряд других признаков сходства с птицами. Полагают, что в эволюции позвоночных было несколько попыток выйти в воздушную среду, некоторые из которых оказались тупиковыми (например Archeopteryx). Более того, одна из переходных от рептилий к птицам форм, названная Protoavis, имея возраст более чем на 75 млн. лет превышающий возраст археоптерикса, обладает большим набором «птичьих» признаков. Расхождение основных стволов птиц на килевые и бескилевые произошло очень рано – в мелу, тогда же началось интенсивное формирование стволов надотрядного и отрядного рангов. Особый интерес к изучению организации ЦНС птиц возник в связи с работами Л.В. Крушинского (сотрудника Московского государственного университета) по изучению рассудочной деятельности у птиц и других позвоночных. Было показано, что головной мозг птиц, особенно врановых, отличается очень высоким уровнем функциональной и структурной организации, а по уровню рассудочной деятельности многие птицы не уступают млекопитающим. У птиц сильно развиты полушария головного мозга и зрительные доли среднего мозга (рис. 10). |
|
Млекопитающие
|
Два главных ствола этого класса сформировались еще на «рептилийной» стадии и ныне представлены первозверями (подкласс Prototheria) и зверями (подкласс Theria). Последние в наше время включают сумчатых (инфракласс Metatheria) и плацентарных (инфракласс Eutheria) млекопитающих. Современные первозвери – утконосые и ехидновые разошлись в своем эволюционном развитии очень рано еще в мезозое. Они крайне уклонились по образу жизни от своих предков и наряду с сохранением древних примитивных признаков приобрели множество специфических адаптаций. Сумчатые и плацентарные представляют собой две ветви млекопитающих, отошедшие от общей предковой группы триботерий (Tribotheria) на рубеже раннего и позднего мела. Консервативными реликтами того времени среди сумчатых являются опоссумовые. Наиболее архаичные плацентарные относились к отряду ныне вымерших млекопитающих из отряда Protheutheria, представители которого жили в конце раннего мела в палеогене. От самых ранних форм протеутерий отделились будущие неполнозубые млекопитающие и (не менее чем тремя параллельными ветвями) формы тенрекообразных, ежеобразных и землеройкообразных, объединяемые в отряд насекомоядных (Insectivora). На этом этапе развития ЦНС у млекопитающих в полушариях конечного мозга происходит значительное увеличение площади неокортекса, появляются ассоциативные зоны. Значительно дифференцируется таламус и ядра конечного мозга. Предполагается, что у насекомоядных и грызунов в организации сложных форм поведения решающая роль принадлежит стриатуму, поэтому этот уровень интеграции ЦНС получил название стриато-кортикальный и представляет собой четвертый критический этап в эволюции ЦНС (рис. 3д). В конце мела именно от протеутерий отделились ветви, давшие первых приматов, рукокрылых и шерстокрылых. Группы протеутерий, давшие начало грызунам, по-видимому стояли уже далеко от тех форм, которые были родоначальниками современных зайцеобразных. Одной из важнейших групп плацентарных, возникших от протеутерий, были древние копытные (отряд Condylarthra), сформировавшиеся в середине позднего мела. Освоив растительный корм они смогли существенно повысить численность, широко распространиться по суше и дать обширную адаптивную радиацию. Из группы древних копытных развились такие ветви как отряды трубкозубых, даманов, сирен, хоботных, парнокопытных, мозоленогих, непарнокопытных, китообразных и др. При этом даже в пределах отряда разные группы могли возникнуть от разных предковых форм (например, зубатые и усатые киты). Становление крупнейшего отряда плацентарных – хищных (Carnivora) шло от протеутерий или от очень ранних древних копытных при переходе последних к плотоядности. По общей организации определенную близость к своим палеогенным предкам имеют псовые и виверровые, тогда как кошачьи являются наиболее видоизмененными и специализированными по сравнению с предковыми формами. Древние хищные, приспособившись к обитанию в воде, смогли дать начало группе ластоногих, среди которых ушастые тюлени, видимо, наименее изменились по сравнению с ранними формами ластоногих, живших на рубеже палеогена и неогена. В отрядах высших млекопитающих (хищных, приматов, хоботных и др.) тенденция к теленцефализации мозга достигла своего максимального развития. Резко увеличился общий размер полушарий и площадь неокортекса, усложнилась его внутренняя структура. Развились сложные формы нервной деятельности, включая рассудочную деятельность и мышление (у человека). Эту стадию развития ЦНС А.И.Карамян рассматривал как высший – пятый критический этап эволюции мозга. Высшие центры интеграции сосредоточиваются в филогенетически молодых и активно развивающихся зонах неокортекса – лобной и теменной коре. В эволюции ЦНС формируется неокортикальный уровень интеграции (рис. 3е). |
|
Заключение |
Разделение головного мозга на пять отделов, происходящих из мозговых пузырей характерно для представителей всех групп позвоночных от круглоротых до млекопитающих и человека. При этом в процессе эволюции структура и функциональная организация этих отделов претерпевают значительные изменения. Полученные к настоящему времени данные заставляют отказаться от традиционных линейных схем эволюции нервной системы по типу «от низших к высшим». В каждой группе современных позвоночных имеются различ¬ные варианты организации нервных центров и мозга в целом, развивающиеся параллельно и независимо друг от друга. В этом плане даже мозг человека представляет собой только один из возможных и реализованных в эволюции вариантов организации ЦНС. Эволюция основных групп позвоночных во многих случаях шла не только независимыми, параллельными рядами, но и неравномерно по времени и скорости. Такой «мозаичный» характер эволюции животных без сомнения отразился и на строении их нервной системы. Важными задачами эволюционной нейроморфологии является выяснение путей эволюционного развития нервной системы и мозга, в частности, факторов, влияющих на их развитие и становление, а также выявление аналогий/гомологий в отделах нервной системы у разных видов животных. Все это является необходимой основой для правильной интерпретации данных, получаемых в различного рода нейробиологических исследованиях представителей позвоночных животных, многие из которых используются в качестве «модельных» объектов для решения целого ряда проблем, стоящих перед физиологическими науками, медициной, токсикологией, биологией развития и т.д. |
|
Рекомендуемая литература |
|
|