
|

|




|
|
|
|
|
|
|
|

|
|
Glischrochilus (Librodor) |
Семейство блестянки – Nitidulidae Latreille, 1807
Число видов в мире и России.
Число описанных к настоящему времени блестянок превысило 3000 видов, объединенных в 271 род и подроды. Однако в мировой фауне можно ожидать по крайней мере 7000 - 8000 видов Nitidulidae, причем на Индо–Малайскую область приходится не менее 2000 видов. Фауна России и стран СНГ насчитывает около 500.
Общая популярная характеристика семейства
и его значение.
Пронырливые жуки – блестянки - могут Вам встретиться где угодно, хотя скорее всего Вы их не заметите – уж очень они малы, всего каких–то 2 – 3 мм или и того меньше. Но если уж задаться целью и попытаться найти этих крошечных существ, то нужно сперва вооружиться лупой, а потом смело идти на ближайший луг, в поле или сад… Осторожно подойдите к какому–нибудь аппетитному с точки зрения жука цветку, присмотритесь – и вот они - наши блестянки – копашатся, толкаются, пытаются залезть внутрь цветка. И так всю их короткую жизнь, день за днем – с цветка на цветок в поисках лакомой пыльцы. Личинки тоже развиваются внутри цветочных бутонов и спускаются на грешную землю только для окукливания.
Можно подумать, что все блестянки живут на цветах. Да, и некоторые из них могут серьезно вредить сельскохозяйственным и декоративным растениям, выедая бутоны и пыльники. Но немало их живет под корой, где хищничают на личинок вредных короедов и долгоносиков. Другие любят полакомиться сочными плодами, а часть представителей, наоборот, освоила продовольственные склады, поедая зерно, сухие фрукты и т.п. и активно расселяясь с помощью человека по всему миру.
Диапазон размеров тела.
Семейство жуков – блестянок объединяет мелких и средних насекомых, размер которых колеблется в пределах 0,8 (подсемейство Cybocephalinae) – 15 мм и более.
Форма тела.
Тело разнообразной формы, обычно более или менее овальное или удлиненное, почти параллельностороннее или даже полуцилиндрическое. Чаще умеренно выпуклое дорсально и уплощено или слабо выпуклое с вентральной стороны. Некоторые имеют сильно выпуклое сверху и уплощенное снизу тело. Для многих блестянок характерна тенденция к укорочению надкрылий и уплощению тела, особенно ярко выраженная среди подсемейств Carpophilinae и Cillaeinae, вследствие чего некоторые циллеины первоначально были описаны в составе семейства коротконадкрылых (Staphylinidae). Паразитоиды кокцид и белокрылок из подсемейства Cybocephalinae имеют во большинстве случаев полусферическую форму тела и способны сворачиваться в шар.
Пунктировка.
Обычно поверхность тела равномерно пунктирована. Иногда помимо пунктировки, заметны различные по размерам вдавления или борозды. Надкрылья иногда несут бороздки, кили и ряды, образованными правильной или неправильной пунктировкой.
Опушение.
Опушение чаще умеренно густое и тонкое, иногда совершенно редуцировано или состоит из групп волосков разного размера, окраски и формы. Некоторые группы подсемейств Nitidulinae и Cillaeinae имеют весьма замысловатые комбинации групп или рядов щетинок и волосков, а мирмекофильные Lawrencerosini несут характерные и для других мирмекофилов трихомы. Нередко формы с богатым опушением имеют краевые реснитчатые ряды по бокам переднеспинки и надкрылий.
Внешний вид.
Голова частично втянута в переднегрудь, прогнатная; усики обычно 11–члениковые с 3–члениковой компактной булавой (очень редко 9-10-члениковые и\или с 1-2 члениковой булавой, а иногда булава становится 4-7 члениковой). Надкрылья у некоторых форм могут быть сильно укорочены, с хорошо развитыми эпиплеврами. Формула лапок 5–5–5, а у Cybocephalinae 4–4–4.
Окраска тела.
Для большинства представителей семейства характерна однотонная окраска тела с преобладанием темных тонов. Имаго подсемейств Epuraeinae, Amphicrossinae и Meligethinae в большинстве случаев характеризуются черной, коричневой, реже рыжей или светло–желтой окраской. Некоторые виды как указанных, так и других подсемейств окрашены в зеленые, синие цвета с металлическим блеском.
Блестянки подсемейства Nitidulinae, наоборот, более разнообразны по окраске и нередко имеют рисунок темных или светлых пятен. Так, тропические формы различных родов (особенно трибы Strongylini) имеют довольно замысловатый орнамент, хотя чаще всего он представлен симметричными пятнами на надкрыльях и переднеспинке. Для палеарктических представитей мицетофильных родов Cychramus Cugelann, 1794, Cyllodes Erichson, 1843, Pocadius Erichson, 1843, Thalycra Erichson, 1843 характерна рыжая или коричневая цветовая гамма, а уже у некрофильных блестянок из родов Omosita Erichson, 1843 и Nitidula Fabricius, 1775 доминируют желтые и светло–коричневые тона, часто образующие на надкрыльях пятна и перевязи. Интересными являются блестянки из рода Soronia Erichson, 1843, которые встречаются на натеках древесного сока совместно с различными представителями Epuraea Erichson, 1834, Amphicrossus Erichson, 1843, Cryptarcha Shuckard, 1839, некоторые из которых имеют леопардовую окраску, что позволяет им великолепно маскироваться на древесной коре.
В подсемействе Cryptarchinae наиболее заметно окрашены жуки рода Glischrochilus Reitter, 1873 - на черных, темно-коричневых надкрыльях обычно располагаются две пары, варьирующих по цвету от желтого до красного, пятен или перевязей. Блестянки рода Cryptarcha, как и Soronia, нередко имеют четко оформленные перевязи и также прекрасно маскируются на коре. Представители рода Pityophagus Shuckard, 1839 имеют темно – коричневую окраску.
Голова.
Голова в большинстве случаев поперечная и отчетливо прогнатная, частично втянута в переднегрудь, хотя иногда (особенно у форм с редуцированной переднегрудью) может сильно подгибаться книзу. Гуларные швы выражены или полностью сросшиеся. Почти во всех случаях на нижней поверхности развиты усиковые бороздки, иногда также заметные постокулярные ямки или дополнительные парамедиальные бороздки.
Лабрум.
Лабрум чаще представлен двумя лопастями или несет небольшую медиальную вырезку, реже лабрум цельный или слит с внутренней поверхностью эпикраниума (подсемейство Cryptarchinae).
Мандибулы.
Мандибулы нередко с заостренными 2-3-зубыми вершинами, обычно с хорошо развитыми молой и простекой.


Мандибула и максилла Carpophilus (Ecnomorphus) lepidus Murray, 1864 (Индонезия, коллекция Музея естественной истории в Лондоне, паралектотип)
Максиллы.
Максиллы во всех группах однолопастные (с лацинией) или с небольшим и плотно прижатым придатком (галеей), с 5-члениковыми щупиками.
Лабиум.
Ментум весьма разнообразен по форме и размерам, чаще попечный, с округлыми очертаниями и медиальный выемкой для лабиума. Лабиум часто простой с мембранозной вершинной частью и трехчлениковыми щупиками.
 |
|
Thalycrodes australe, |
 |
|
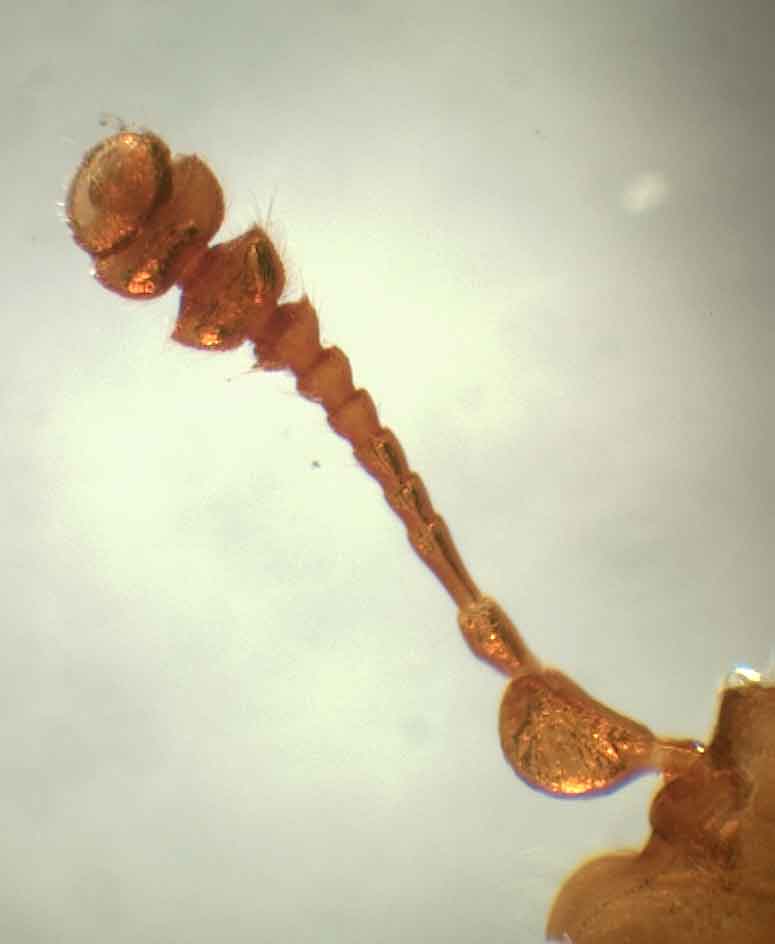
Усик Carpophilus (Ecnomorphus) |
Усики.
Усики чаще всего 11–члениковые, с 3-4–члениковой, компактной и более или менее дорсовентрально уплощенной булавой, хотя у большинства представителей подсемейства Epuraeinae булава только слабо уплощена. Иногда усики состоят из 10 или меньшего количества члеников, и тогда число члеников в булаве сокращается до 2 или включает еще один дополнительный членик – вплоть до 5 – 8 (в этом случае усики имеют полную компактную булаву или частично редуцированную). Однако, виды рода Calonecrus J. Thomson, 1857 обладают 10-члениковыми усиками с 1-члениковой не уплощенной булавой.
Глаза.
Глаза хорошо развиты, особенно крупные у открытоживущих форм (чаще всего у антофильных и паразитоидов равнокрылых), обычно полусферической формы и сильно выступающие у многих представителей семейства. У видов подсемейства Cryptarchinae и других скрытоживущих форм глаза сильно смещены на бока головы (особенно в роде Pityophagus Shuckard, 1839), а иногда также заметно уплощены.
Переднегрудь и переднеспинка.
Переднеспинка сильно изменчива по очертаниям, но всегда с резким перегибом по боковым краям. Отросток переднегруди почти во всех случаях хорошо развит и принимает важное участие в сочленении передне- и среднегрудного сегментов. Передние тазиковые впадины чаще не полностью замкнутые или замкнутые сзади за счет гипомер или, примыкающих или сросшихся с вершиной отростка переднегруди, а в случаях его редукции сросшихся медиально и образующих единую медиальную полоску. Только у некоторых Strongylini, Cychramptodini и Lawrencerosini (Nitidulinae), а также всех Cybocephalinae переднегрудь сильно редуцирована.
Среднегрудь и щиток.
Среднегрудь, как правило, вдавлена у заднего края переднегруди. Щиток маленький, часто почти треугольный или четырехугольный и с закругленной вершиной.
Заднегрудь.
Заднегрудь всегда крупная, нередко с частично выраженным медиальным швом, вдоль заднего края средних тазиковых впадин обычно заметны затазиковые линии.
Ноги.
Все тазики блестянок поперечные: передние и средние – слабовыпуклые, с отчетливо выступающим трохантином, а задние – уплощенные и менее подвижные. Вертлуги промежуточного типа между нормальным и гетеромероидным и короткие. Передние голени обычно треугольные, с наружным кантом, несущим ряд зубчиков, между которыми иногда находятся сенсиллы, а на вершине заметны две не равной длины шпоры; для многих Meligethinae характерно значительное развитие зубцов и коротких тангорецепторов между ними. Средние и задние голени не сильно уплощены, по наружному краю с двумя продольными кантами, вооруженными щетинковидными сенсиллами, но для некоторых форм иногда характерны широкие и плоские голени, а один или оба наружных канта редуцируется. Бедра имеют выемки для сочленения с основанием голени. Лапки 5-члениковые (у Cybocephalinae – 4–члениковые), при этом лапка архаичного типа состоит из трех 2–лопастных члеников с подушечками из волосков, четвертого – самого маленького, иногда плохо заметного между лопастей третьего, и пятого, самого длинного членика с 2 коготками, между которыми иногда заметен рудимент двущетинкового эмподия.
Крылья.
Крылья с сильно редуцированным жилкованием, самое большое с тремя анальными жилками. Радиальные и анальные ячейки, а также и субкубитальное пятно отсутствуют.
Надкрылья.
Надкрылья чрезвычайно изменчивы, с четко окаймленными сторонами и обычно более или менее уплощенные и часто неполные (укороченные), причем наиболее укороченные у видов подсемейства Cillaeinae. Эпиплевры развиты, с резким перегибом к вентральной поверхности, в большинстве случаев они приподняты кнаружи (особенно у форм с более укороченными элитрами), хотя у ряда форм с выпуклой формой тела они резко ниспадают.
Брюшко.
Брюшко состоит из 5 подвижно сочлененных вентритов и 7 тергитов. Вдавление, проходящее вдоль основания пигидия, может преобразовываться в 1-5 пар коротких, но глубоких полулунных вдавлений (комплекс Aethina из Nitidulinae), а у всех представителей Meligethinae эти структуры представлены только парой довольно глубоких и широких вдавлений. Гипопигидий обычно простой, но у большинства Meligethinae его основание с парой вдавлений, сходных с таковыми на пигидии представителей этой группы. Медиальная часть гипопигидия у видов Amphicrossinae обособлена по бокам мембранизованными полосками и превратилась в подвижную лопасть, а у всех Carpophilinae гипопигидий глубоко вырезан у вершины.
Дыхальца.

Дыхальца вершины брюшка Carpophilus (Carpophilus) flavipes Murray, 1864
Формы с полными элитрами имеют маленькие, овальные и одинаковые брюшные дыхальца, расположенные обычно между тергитами и латеростернитами 1–6 сегментов. Большинство форм с укороченными элитрами обладают большими дыхальцами, находящимися на сегментах перед вершиной брюшка (довольно часто сильно поперечные и расположены вдоль оснований тергитов). Дыхальца на непокрытых сегментах обычно очень маленькие и едва заметны.
Эдеагус.
Эдеагус представлен 2 типами.
Карпофилинный тип: тегмен (латеральные доли) состоит из двух лопастей с более или менее глубокой срединной выемкой, а ствол пениса (медиальная лопасть) мембранозный и слабо уплощенный или неуплощенный (Epuraeinae, Carpophilinae, Amphicrossinae, Calonecrinae);
Нитидулинный тип: тегмен состоит из одной пластинки или с короткой медиальной выемкой, а ствол пениса обычно сильно склеротизован и дорсовентрально уплощен (Maynipeplinae, Nitidulinae, Meligethinae, Cillaeinae, Cryptarchinae, Cybocephalinae).
Эдеагус неинвертированного кукуйоидного типа. Эдеагус блестянок карпофилинной линии сходен в соотносительном расположении и форме склеритов с эдеагусами трехлопастного типа элатериформных и ложнотрехлопастного типа тенебриноидных. Нитидулинный эдеагус более простого строения и принципиально отличного функционирования, его тегмен и ствол пениса имеют вид плоских пластинок, хотя очертания обоих склеритов в различных группах этой линии обнаруживают более широкий диапазон изменчивости в сравнении с их гомологами у групп карпофилинной линии. Функционирование эдеагусов обоих типов значительно отличается. Ствол пениса и его внутренний мешок карпофилинного типа в выпяченном наружу эдеагусе проходит между лопастями тегмена, тогда как нитидулинный эдеагус в выпяченном наружу виде раскрывается двумя створками, а внутренний мешок выбрасывается из вершинного пениса.
Анальный склерит (? 8-й тергит) самца вместе с базальной (вентральной) пластинкой и примыкающей к ней непарной гастральной спикулой (spiculum gastrale) образуют характерную для блестянок и близких к ним семейств генитальную капсулу.
Гениталии самки не обнаруживают значительных модификаций от генерализованного кукуйоидного типа, при этом масштаб их изменчивости немного меньше, чем тот, который наблюдается в строении эдеагуса. Как правило, парные гонококситы состоят из внутренних и наружных долей, а на вершине они несут короткие стили. Под яйцекладом находится вентральная спикула (spiculum ventrale), участвующая в образовании генитальной капсулы.

Механизм работы эдеагусов. Сагиттальный разрез (по Kirejtshuk, 1986)
Calonecrinae, Epuraeinae, Carpophilinae, Amphicrossinae:
а – эдеагус в покое,
b – то же с вывернутым внутренним мешком пениса;
Meligethinae, Nitidulinae, Maynipeplinae, Cillaeinae, Cryptarchinae, Cybocephalinae:
c – эдеагус в покое, d - то же с вывернутым внутренним мешком пениса
(с – анальный склерит, т – тегмен, п – пенис)
Половой диморфизм.
Половой диморфизм среди представителей семейства Nitidulidae выражен в различной мере: и довольно слабой, и очень значительной. Наиболее характерным типом диморфизма являются отличия в строении склеритов последнего сегмента брюшка, в частности в форме вершин пигидия и гипопигидия. Выступающий наружу анальный склерит у самцов всех Epuraeinae, Carpopohilinae, Calonecrinae, Maynipeplinae и Cybocephalinae, многих Cillaeinae, а также некоторых Nitidulinae и Cryptarchinae нередко сопряжен с измененным очертанием вершины пигидия, а у всех Carpophilinae и Amphicrossinae резкие отличия наблюдаются также в строении гипопигидия. Однако самцы многих палеарктических блестянок имеют на гипопигидии небольшие отличия в очертании заднего края, дополнительные бугорки, небольшие кили или блестящие площадки у вершины.
У блестянок из палеарктического рода Pria Stephens, 1829, как и у многих других родов Meligethinae, эндемичных в субтропических и тропических территорий, самцы имеют более длинные усики с булавой, включающей большее число члеников, чем булава самки, почти всегда состоящая из 3 члеников. Такой диморфизм характерен для большинства Mystropini, а также для отдельных групп других триб Nitidulinae, некоторых Epuraeinae и Carpophilinae.
Помимо указанных типов диморфизма, самцы и самки могут иметь различия в размерах головы, мандибул и переднеспинки, а также в форме и строении эпикраниума, лабрума, отростка переднегруди, заднегруди и ног (наиболее обычны более широкие передние лапки у самцов).
Яйца.
Яйцо мягкое, удлиненно – овальной формы, без рельефной микроскульптуры.
Личинки.
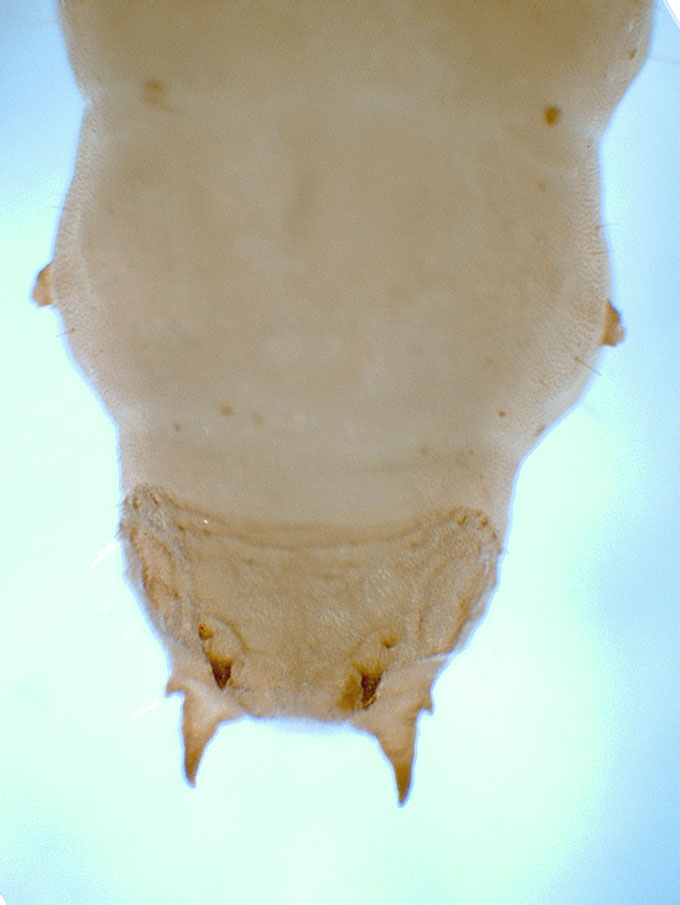
Зрелые личинки от 1 до 20 мм в длину. Тело обычно овальное в поперечном разрезе или несколько уплощенное дорсовентрально, слабо склеротизованное, однако эпикраниум с придатками, ноги, пластинка переднеспинки и небольшие участки на других тергитах сильно склеротизованные. Спина гладкая или часто с бугорками, отростками, возвышениями или выступами различной конфигурации, иногда более или менее склеротизованными. 9-й брюшной тергит с прегомфами и урогомфами (особенно развитыми у форм, живущие внутри субстрата, и редуцированными – у свободноживущих, отсутствующими у Cybocephalinae).
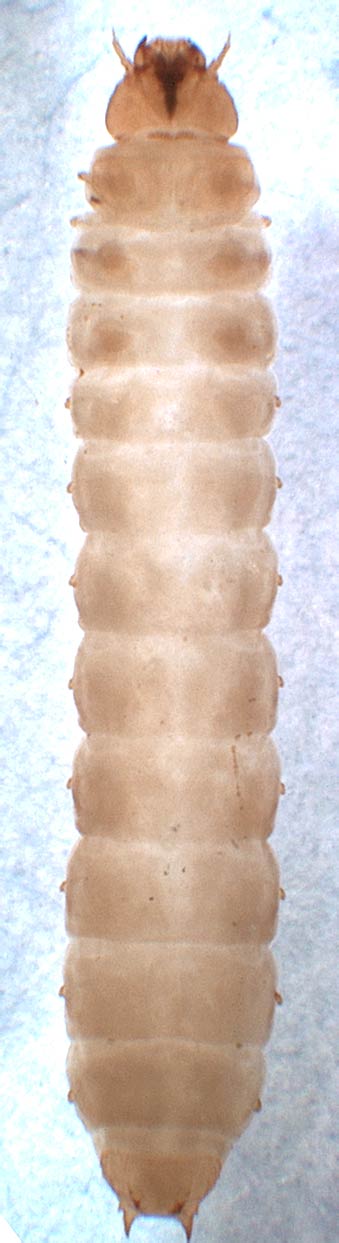

Личинка Carpophilus (Megacarpolus) titanus Murray, 1864 (Приморский край, под корой, в бродящем соке березы)
Голова прогнатная, со лбом, слитым с наличником, но с развитым фронтоклипеальным швом, 2–4 глазками на каждой стороне (если видны) и отсутствующей эндокариной; гипостомальные складки обычно сильно расходятся кзади; лаброклипеальный эпифаринкс имеет медиальный шов и множество маленьких складок, отсутствующих у некоторых видов – антофагов из Epuraeinae, Meligethinae и Cybocephalinae, а также с более или менее слабо склеротизированными грудными и брюшными сегментами. Фронтальный шов сзади доходит до заднего края головной капсулы, образуя вдоль него поперечную линию, только у видов Pocadius базальная часть шва превращена в медиальную линию, а у видов Cillaeinae она имеет вид острого угла. Медиальная эндокрина отсутствует. Усики 3-члениковые. Мандибулы большей частью хорошо развиты, с простекой и отчетливо вооруженной молярной частью, но у личинок представителей Meligethinae простека представлена лишь мембранозным выростом, а у видов Mystropini и Cybocephalinae простека и мола полностью редуцированы. Максиллы однолопастные, с мала (слившиеся лациния и галеа), с большим количеством микротрихий и обычно с развитым пальцевидным придатком, несущим 3-члениковые щупики, однако у антофильных Mytropini и Meligethinae мала без выраженного опушения или микротрихий, а у видов Cybocephalinae она и без заметного пальцевидного придатка; кардины явные; гипофаринкс со склеромом и браконами (исключая Cybocephalinae). Лабиум в значительной мере редуцирован, с 1-члениковыми щупиками.
 |
 |
 |
|
Meligethes schilskyi Rtt., 1896;
личинка старшего возраста:
|
||
Дыхальца двугубные и расположены на вершине дыхальцевой трубочки (если присутствует) у личинок, обитающих в жидких субстратах; брюшные дыхальца расположены дорсолатерально на 1-8-ом сегментах или редко под боковыми расширениями каждого сегмента (некоторые Cybocephalinae имеют кольцевые дыхальца с 2 боковыми воздушными трубочками). Ноги скорее короткие с редкими, тонким и заостренными щетинками; тарзунгулюс свободноживущих личинок иногда с субапикальным сенсиллярным пузырьком или головчатой щетинкой (Meligethinae; Nitidulinae, Mystropini и Cybocephalinae). 9-й сегмент брюшка обычно несет простые урогомфы и нередко – прегомфы.
Куколки.
Форма куколок находится в значительной корреляции с формой тела имаго. Голова всегда подогнута на вентральную сторону, полностью покрыта передней частью переднеспинки имеет несколько надглазничных бугорков, несущих тонкие, длинные и заостренные щетинки; лоб обычно с обычно обособленным и выпуклым наличником. Переднеспинка с парой длинных острых выростов, расположенных ближе к переднему краю, или с несколькими (2–3) парами выростов с вершинной щетинкой, расположенной апикально, субапикально или базально, а в некоторых случаях выросты редуцированы или вытеснены сравнительно длинными щетинками. Среднеспинка, надкрылья и заднеспинка обычно гладкие; медиальная часть заднего края средне- и заднеспинки выступает дуговидно или угловидно. Заднее бедро и довольно часто среднее несут 1–3 субапикальные щетинки. Каждый брюшной тергит гладкий или часто тергиты I–IV имеют пару парамедиальных выростов, хотя каждый латеростернит (I–VIII) снабжен 1–2 парами щетинконосных бугорков или щетинками, расположенными между дыхальцами и латеральным краем (обычно дыхальца имеют по одной щетинке). Первые 7 сегментов всегда хорошо развиты. 8-й и 9-й – в различной степени втянуты внутрь, а вершина брюшка с парой длинных простых “урогомф” (даже у видов, чьи личинки не имеют урогомф вообще – а именно: Meligethinae и Cybocephalinae).



Куколка Carpophilus (Megacarpolus) titanus Murray, 1864 (Приморский край, под корой, в бродящем соке березы)
Диагноз и систематическое положение.
Семейство Nitidulidae относится к надсемейству Cucujoidea и, по-видимому, имеет наиболее близкие филогенетические связи с семейством Helotidae, хотя нередко его сближали с семействами Katerertidae, Monotomidae (=Rhizophagidae) и Smicripidae (Crowson, 1955; Кирейчук, 1982, 1986; Lawrence, 1982, 1991; Audisio, 1993 и Kirejtshuk, 1998, 2000).
Имаго блестянок характеризуются формулой лапок 5-5-5 у обоих полов (у представителей Cybocephalinae – 4-4-4), отсутствием дыхалец не только на 8-м, но и на 7-м сегментах брюшка, хорошо выраженной булавой усиков, кольцевым основанием тегмена за счет причлененного вилковидного склерита. Облик некоторых блестянок, особенно представителей комплекса родов, близких к Soronia, довольно похож на представителей семейства Peltidae. Кроме того, некоторые другие имагинальные признаки блестянок весьма генерализованы (по-видимому, в большинстве случаев первично), а именно: поперечные передние и средние тазики с выступающим наружу трохантином, 5 видимых вентритов, мандибулы с молой и простекой, незаросшие гуларные швы, а у личинок таковыми могут рассматриваться обычно развитые немодифицированные урогомфы. Вместе с тем, другие признаки свидетельствуют о значительной продвинутости группы в целом - у имаго: отстствие дыхалец на 7-м сегменте брюшка, весьма редуцированнное жилкование задних крыльев с проксимальным смещением, выраженная тенденция к коротконадкрылости и однолопастные максиллы, а у личинок: полностью сросшийся лабрум со лбом, 1-члениковые лабиальные щупики и 8 пар брюшных дыхалец.
В отличие от сходных с ними представителей семейства Kateretidae, взрослые блестянки имеют максиллу, несущую только широкую лацинию, хотя у некоторых к ней плотно примыкает очень узкая и маленькая галея, сравнительно небольшой 6 сегмент брюшка, нередко негомономные дыхальца брюшка; а также симметричное строение эдеагуса без причленяющихся парамер; в то время как их личинки отличаются от Kateretidae отсутствием эндокарины в эпикраниуме, округло сходящимися гипостомальными гребнями, отсутствием гипостомальных утолщений (за исключением некоторых Cybocephalus, у которых они выражены).
От сходных с ними представителей семейства Smicripidae взрослые блестянки отличаются отсутствием фронтоклипеального шва, 3-члениковыми лабиальными щупиками, более или менее редуцированными нотостернальными швами, умеренно длинным последним сегментом брюшка, симметричным строением эдеагуса без причленяющихся парамер, а личинки – отчетливым базальным члеником максилл (кардо) и отсутствием или расходящимися гипостомальными утолщениями.
 |
|
Maynipeplus lomechusoides |
От представителей семейства Monotomidae взрослые блестянки отличаются, как правило, 11-члениковыми усиками чаще с 3-члениковой булавой, однолопастной максиллой, незамкнутыми или неполностью замкнутыми сзади передними тазиковыми впадинами, поперечнми передними и средкими коксами с выступающим трохантином, формулой лапки 5-5-5 или 4-4-4 у обоих полов, симметричным эдеагусом без причленяющихся парамер и с одной аподемой ствола пениса, а личинки округло сходящимися гипостомальными гребнями, длинным базальным члеником максилл (кардо), обычно отсутствущимися гипостомальными утолщениями длинными двухчлениковыми лабиальными щупиками, неразветвленными урогомфами.
От родственных им представителей семейства Helotidae, взрослые блестянки отличаются неудлиненными глазами, однолопастными максиллами, отсутствием отчетливых субментальных бороздок (? гомологи или аналоги которых, однако, заметны у видов трибы Mystropini), незамкнутыми или неполностью замкнутыми сзади передними тазиковыми впадинами, поперечнми передними и средкими коксами с выступающим трохантином, редукцией и проксимальным смещением жилок задних крыльев, отсцутствием дыхалец на 7-м сегменте брюшка, склеротизованной гастральной спикулой и непарной аподемой пениса, а их личинки – округло сходящимися гипостомальными гребнями, обычно более длинным базальным члеником максилл (кардо), одночлениковыми лабиальными щупиками, неразделенными ментумом и субментумом, сравнительно короткими или редуцированными урогомфами.
Развитая вентральная пластинка, слитая или сочлененная с гастральной спикулой (spiculum gastrale) у самцов вместе с анальным склеритом (вероятно, производным рудиментом 8–го тергита и\или частично 8–х латеростернитов) образуют генитальную капсулу, характерную для представителей различных семейств (Nitidulidae, Brachypteridae, Smicripidae). Сходные образования самок формируются из вентральной спикулы (spiculum ventrale), обычно рассматривающейся дериватом 9-го брюшного сегмента. Также в этих семействах наблюдается тенденция к редукции имагинальной галеа (как в семействе Monotomidae, в то время как многие Polyphaga - такие как представители Boganiidae (Cucujoidea), Chrysomeloidea, Curculionoidea – чаще имеют тенденцию к редукции лацинии); отсутствуют функционирующие дыхальца на 7- брюшном сегменте у имаго; отсутствуют латеральные расширения (пластинки) имагинального метэндостернита (обычного у архаичных Cucujoidea, а также и у Cleroidea, Lymexyloidea, Tenebrionoidea, Chrysomeloidea и Curculionoidea); членистая максиллярная мала (?=галеа) со склеротизованным придатком (?=лациния) у личинок) (как во многих случаях среди Boganiiae, Helotidae, некоторых Cleroidea, Chrysomeloidea и Curculionoidea, но иногда и среди Lyctidae, Ptinidae, Peltidae, Lymexylidae, Endomychidae); 1–члениковые лабиальные щупики у личинок (также встречаются среди Lyctidae, Anobiidae и некоторых Monotomidae); личиночный эпикраниум не более чем с 4 стеммами (простыми глазками) (обычно 2 или 4) на каждой стороне (хотя данная особенность также не редка и среди личинок Polyphaga).
Имеются некоторые свидетельства (палеонтологические, морфологические, экологические и биономические) для предположения о том, что формирование генерализованных Epuraeinae, Carpophilinae, Meligethinae и других подсемейств, в отличие от мнения Crowson (1988), скорее связано с более быстрым онтогенезом (акселерацией развития), нежели чем с архаичным характером некоторых их структур.
Можно предположить следующие признаки для ближайших предков блестянок:
Имаго: овальное тело, слегка выпуклое дорсально и вентрально, с пунктировкой и опушением подобным таковому у представителей родов Soronia Erichson, 1843 (Nitidulidae), Ericmodes Reitter, 1877/1878 (Protocucujidae), Lophocateretes Olliff, 1883, Zimioma des Gozis, 1886, Ostoma Laicharting, 1781 и Thymalus Latreille, 1802 (Peltidae); прогнатическая голова с двухлопастным свободным лабрумом, выраженной мандибулярной молой и простекой, однолопастной максиллой; широко уплощенными сторонами переднеспинки и элитр; элитры полные, с широкими и полными эпиплеврами; передние тазики разделены сравнительно широким отростком переднегруди, который далеко выдается в виде складки на поверхность среднегруди; среднегрудь с хорошо развитыми медиальными и паракоксальными швами; поперечные метакоксы, которые не наклонены медиально как у Cleroidea (включая вероятных including Peltidae); все вертлугти кукуйоидного (нормального) или почти тенебрионоидного (гетеромерного) типов; передние голени с наружным зубчатым краем, а средние и задние с 2 кантами, несущими щетинки вдоль наружнего края; вершины всех голеней с парой шпор; лапки с тремя 2-лопастными члениками, 5-й членик - самый длинный и оканчивается хорошо заметным двущетинковым эмподием между коготками; анальный склерит полностью втянут в 8-ой брюшной сегмент у обоих полов; вентральная пластинка у самцов разделена на 2 части, соединенных гастральной спикулой; слегка склеротизованный тегмен окружает слабо склеротизованный и уплощенный дорсовентрально ствол пениса (как у представителей родов Axyra Erichson, 1843, Megauchenia Macleay, 1825, Prometopia Erichson, 1843, Platychora Erichson, 1843 и др.); вилковидный склерит соединен с тегменом; ствол пениса с непарной аподермой у основания и парными лопастями, закрывающими субапикальное отверстие.
Личинка: тело удлиненное, слабо склеротизованное, исключая сильно склеротизованный эпикраниум с придатками, пластинку переднеспинки и небольшие участки на других тергитах; голова и лоб слиты с наличником; лаброклипеальный эпифаринкс несет медиальную складку и множество небольших линий; мандибулы с развитой молой и простекой; максиллы с мала и 3–члениковыми щупиками; лабиальные щупики 1–члениковые; гипофаринкс со склеромом и браконами; дыхальца располагаются на вершине дыхальцевых трубочек.
Местообитания и биономия
Представители семейства Nitidulidae встречаются во многих типах биотопов, но почти отсутствуют в сильно увлажненных местах и никогда не встречаются в водоемах. Вместе с тем можно указать, что большинство блестянок почти всех надсемейств так или иначе связано с надземными частями деревьев и лесными стациями. Однако большинство представителей подсемейства Meligethinae, некоторые виды трибы Cychramini (роды Xenosrongylus и Kirejtshukistrongylus Audisio et Jelinek in Audisio, Mariotti, Jelinek et de Biase, 2001: Nitidulinae) и подсемейства Cillaeinae, трофически связанные с цветками преимущественно травянистых растений, нередко приурочены к открытым пространствам, а отдельные специализированные группы подрода Micruria (род Epuraea: Epuraeinae) и некоторых родов триб Nitidulini и Strongylini (Nitidulinae) строго не связаны с надземными частями деревьев и держатся разлагающейся растительной органики в подстилке, у почвы или же в плодовых телах наземных и подземных грибов, причем некоторые из них характерны для открытых пространств. Разнообразие образа жизни в различных группах менее выражено на территориях с умеренным климатом и в высокогорных районах, но становится исключительно широким в регионах с субтропическим и тропическим климатом.
 |
|
Calonecrus wallacei |
Большинство групп рассматриваемого семейства имеет тесные связи с лесными экосистемами, богатыми древесными и кустарниковыми формами растений. Подсемейства Epuraeinae, Amphicrossinae, Calonecrinae и Cryptarchinae - характерные лесные группы, в то время как только большинство представителей Carpophilinae, Nitidulinae и Cillaeinae населяют разные типы лесов и связаны с грибами, развивающимися на тканях деревьев и кустарников, в вытекающем древесном соке и т.п. субстратах. Также довольно обычны в лесных экосистемах и виды родов Pocadius Erichson,1843; Thalycra Erichson, 1843; Quadrifrons Blatchley, 1916; Thalycrodes Blackburn, 1891 из подсемейства Nitidulinae, связанные в питании и развитии с подземными грибами. Даже группы, обитающие на цветах или плодах, часто встречаются на живых или мертвых деревьях и кустарниках [все представители трибы Mystropini, а также многие антофильные формы подсемейств Epuraeinae, Carpophilinae, Meligethinae (Cryptarchopria Jelinek, 1975; Kabakovia Kirejtshuk, 1979; Palmopria Endrody-Younga, 1978 и др.)]. Хищные и паразитические Cybocephalinae и Cychramptodini из подсемейства Nitidulinae в большей степени представлены в экосистемах с деревьями и кустарниками, где обитают колонии кокцид и белокрылок. Наконец, многие виды, развивающиеся на травянистых растениях, предпочитают находиться внутри лесных сообществ (подсемейство Meligethinae и другие). Тем не менее, виды - некрофаги родов Nitidula Fabricius, 1775 и Omosita Erichson, 1843 sensu lato чаще более обычны за пределами лесных экосистем. И только немногие роды могут рассматриваться как характерные для открытых травяных экосистем (таких как многие пустынные и степные группы рода Meligethes Stephens, 1832, sensu lato из подсемейства Meligethinae и кактусоядные Camptodes Erichson, 1843 из подсемейства Nitidulinae).
Многие формы Nitidulidae более регулярно собирается в условиях умеренного и субтропического климата в Северном полушарии с середины апреля до конца мая или очень рано летом (март - июнь), а в Южном полушарии пик активности имаго большинства видов приходится на последние месяцы календарного года. Однако некоторые виды в условиях субтропического и тропического климата скорее более обычны на протяжении всего года, а другие имеют, как правило, интервалы в активности в течение неблагоприятных периодов лета и/или зимы. Низинные дождевые леса населены некоторыми видами блестянок, демонстрирующими круглогодичную имагинальную активность. Обычно эти виды заселяют подкоровые пространства, цветы и мягкие плоды.
Наиболее древними или схожими с ними местообитаниями, вероятно, являются те, которые заселяются неспециализированными грибоядными формами из разных групп семейства Nitidulidae и различных семейств инфраотряда Cucujioformia в целом (Peltidae, Lophocateridae, Phloiophilidae- из надсемейства Cleroidea; Derodontidae и Nosodendridae из надсемейства Dermestoidea; многие семейства из надсемейств Cucujoidea и Tenebrionoidea). В особенности, архаичным образом жизни, возможно, характеризуются блестянки, населяющие вытекающий древесный сок, такие, как виды рода Epuraea sensu lato (Epuraeinae); большинство представителей подсемейств Amphicrossinae и Calonecrinae, рода Soronia (Nitidulinae) и подрода Librodor Reitter, 1884 (род Glischrochilus: Cryptarchinae). Эти особенности, по - видимому, также присущи многим представителям разных подродов рода Epuraea sensu lato (подрод Epuraea sensu stricto; Epuraeanella; Aphenolia Reitter, 1884; Africaraea Kirejtshuk, 1989a; Amedanyraea Kirejtshuk et Pakaluk, 1996 и т.д.) из Epuraeinae;многие или многим представителям подродов Carpophilus Erichson, 1843 sensu stricto и Ecnomorphus Motschulsky, 1858 из подсемейства Carpophilinae; а также большинству видов из трибы Strongylini (исключая виды рода Camptodes Erichson, 1843) из подсемейства Nitidulinae большинству видам трибы Cryptarchini из подсемейства Cryptarchinae. Многие полные мицетофаги из трибы Nitidulini и подсемейства Cillaeinae несут в себе выраженные в большей или меньшей степени элементы архаичного образа жизни и приурочены к соответствующим местообитаниям. Они отчетливо прослеживаются среди многих древесных блестянок из родов Prometopia; Parametopia Reitter, 1884; Lobiopa Erichson, 1843; Axyra; Megauchenia; Ipidia Erichson, 1843; Platychora; Taracta Murray, 1867; Psilotus Fischer, 1829; Perilopa Erichson,1843; Gaulodes Erichson, 1843; Ussuriphia Kirejtshuk, 1992; а также и среди некоторых представителей родов Pocadites Reitter,1884; Hebasculinus Kirejtshuk, 1992; Atarphia Reitter, 1884 и подродов Aethina Erichson, 1843 sensu stricto и Circopes Reitter, 1873, а также подрода Phenolia Erichson, 1843 из трибы Nitidulini. В менее выраженой форме они, возможно, допустимы в особенностях родов Ecnomaeus Erichson, 1843; Cillaeus Castelneau, 1835; Cillaeopsis Grouvelle, 1899; Platynema Ritsema, 1885; Ithyphenes Murray, 1864; и, по крайней мере, часть видов родов Colopterus Erichson, 1842; Brachypeplus Erichson, 1842; Conotelus Erichson, 1843 из подсемейства Cillaeinae. В меньшей степени архаичные признаки выражены среди форм, полностью или частично связанных с подстилкой и разлагающимися субстратами у поверхности почвы или в почве, а иногда и с гниющими плодами, фруктами и семенами. Такие представители есть в подродах Urophorus Murray, 1864 sensu stricto и Anophorus Kirejtshuk, 1990b; родах Stelidota Erichson, 1843; Pocadius; Thalycra; Quadrifrons; Thalycrodes; подродах Lasidites Jelinek, 2001; Aethinodes Blackburn, 1891 и Plesiothina Kirejtshuk, 1990a; а также, возможно, у некоторых видов рода Physoronia Reitter, 1884 из три бы Nitidulini.
Большинству мицетофагов характерно развитие на ферментирующихся дрожжевыми грибами субстратах. Поэтому многие мицетофильные виды могут распространять эту группу грибов. Тем не менее, многие представители различных подсемейств предпочитают или связаны с плодовыми телами базидиомицетов [ такие как некоторые виды подродов Epuraea sensu stricto; Aphenolia; Epuraeanella; Phenolia; Pocadius; Pocadites; Thalycra; Cyllodes; Pallodes Erichson, 1843; Neopallodes; Tricanus Erichson, 1843; Oxycnemus Erichson, 1843]. Некоторые виды подрода Haptoncus, родов Carpophilus, Colopterus, Glischrochilus и других известны как переносчики спор грибов родов Ceratocystis (из Ascomycetes) или Fusarium (из Hyphomycetes).
 |
 |
|
Cylindroramus sp., тело сверху и сбоку (по: Lawrence & Kirejtshuk, 1999)
|
|
Также среди блестянок можно также проследить некоторые закономерные изменения в экологии, трофике и образе жизни, и соответствующие структурные преобразования (Кирейчук, 1989b, 1996b). Наиболее ярко корреляция между экологическими, биономическими и структурными особенностями выражена в группах, которые стали независимо антофагами и используют пыльцу как главный источник пищи (разные группы из подсемейств Epuraeinae, Carpophilinae и Cillaeinae, триб Nitidulini, Strongylini и Cychramini из подсемейства Nitidulinae, а также все представители из трибы Mystropini из Nitidulinae и все представители Meligethinae - Kirejtshuk, 1994a, 1996a). Довольно часто соцветия растений служат временным местообитанием для форм, склонных к фитофагии [например, для некоторых среди представителей подродов Micruria Reitter, 1875a (Hayashi, 1978); Haptoncurina и Parepuraea Jelinek, 1977 рода Epuraea (Kirejtshuk, 1998/1999) из подсемейства Epuraeinae; различных подродов Carpophilus (Connell,1956) из подсемейства Carpophilinae; Neopocadius Grouvelle, 1906c (Bruch, 1923) и Camptodes из подсемейства Nitidulinae; и некоторые Brachypeplus sensu lato (Кирейчук, 1994a, 1996a) из Cillaeinae, хотя другие из упомянутых родственных групп уже стали полными антофагами]. Многие блестянки, прошедшие через эту стадию экологических изменений и ставшие полными антофагами как на личиночной, так и на имагинальной стадиях жизненного цикла, остаются на стадии экологических изменений по сей день [род Propetes sensu lato и подрод Apria Grouvelle, 1919 рода Epuraea (Jelinek, 1992) из Epuraeinae; подроды Caplothorax Kirejtshuk, 1996b и Plapennipolus) Kirejtshuk, 1996b; а также и подрод Urocarpolus Kirejtshuk, 1996b рода Carpophilus, а также род Nitops Murray, 1864 из Carpophilinae; все Meligethinae; подроды Ithyra Reitter, 1873; Idaethina Gemminger et Harold, 1868 (non Reitter, 1875); Сleidorura Kirejtshuk et Lawrence, из трибы Nitidulini (Nitidulinae); возможно, все виды трибы Mystropini (Nitidulinae) и рода Macrostola из подсемейства Cillaeinae]. Следующей стадией этих закономерных изменений в образе жизни и трофике, является переход от антофагии к карпофагии (в подсемействах Epuraeinae и Carpophilinae) или к полной филлофагии (виды родов Anister Grouvelle, 1901; Xenostrongylus Wollaston, 1854; Kirejtshukistrongylus из трибы Cychramini: Nitidulinae).
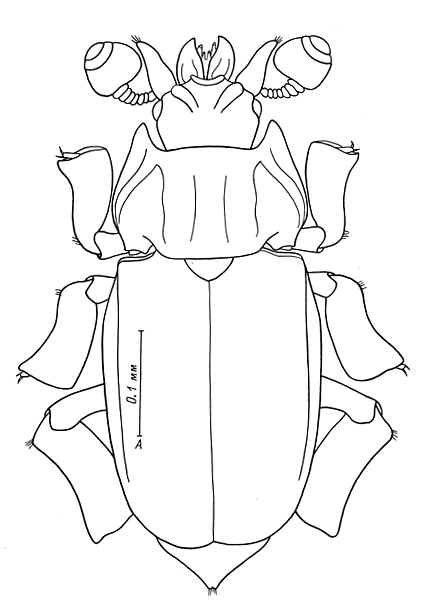 |
|
Krakingus johni Kirejtshuk,
|
Виды родов Nitidula Fabricius, 1775 и Omosita Erichson, 1843 sensu lato связаны с грибами, растущими на трупах позвоночных животных и стали обычными обитателями мусора и сельскохозяйственных и промышленных отходов, содержащих остатки животных. Небольшое число видов рода Epuraea (Epuraea) sensu stricto приурочены к жизни в условиях пустыни и на других аридных территориях, где питаются грибами, растущими на экскрементах млекопитающих. Некоторые мицетофаги Amphicrossus и Amphotis довольно часто имеют симбиотические связи с муравьями (африканский вид Amphicrossus parallelus Grouvelle, 1912 описан из муравейников рода Nitidopecten Reichensperger, 1913; а европейские виды рода Amphotis часто встречаются в гнездах муравьев рода Lasius Fabricius, 1805 - Hymenoptera, Formicidae). Тем не менее, развитие Amphotis marginata (Fabricius, 1781) также связано и с галлами Biorhiza pallida (Olivier, 1791) (Hymenoptera, Cynipidae) (Lengerken, 1941). Представители трибы Lawrencerosini являются облигатными мирмекофилами (Kirejtshuk, 1990c). Некоторые африканские представители подрода Aethina sensu stricto регулярно встречаются в гнездах пчел [A.(A.) tumida Murray, 1867 известен из ульев медоносной пчелы - Lundie, 1940, а родственники собраны из гнезд других пчелиных Apidae (Kirejtshuk, 1998/1999)]. Австралийские виды рода Onicotis Murray, 1864 и некоторые виды из одного из подродов подрода рода Brachypeplus также приурочены к гнездам Apidae.
Некоторые древесные формы, относящиеся к мицетофагам, стали факультативными или облигатными хищниками личинок насекомых и других мелких беспозвоночных, живущих под корой и в древесине, а иногда становятся и постоянными обитателями ходов короедов Scolytinae [некоторые Epuraea (Epuraea) sensu stricto; Glischrochilus (Glischrochilus) Reitter, 1873 sensu stricto; Pityophagus Shuckard, 1839 и другие]. Однако, только в нескольких группах хищничество может рассматриваться как их характерная таксономическая особенность. Хищные формы, охотящиеся на червецов, щитовок, кокцид и белокрылок, есть среди многих видов трибы Cychramptodini из подсемейства Nitidulinae и большинства представителей подсемейства Cybocephalinae (не обнаруживающих следов как близкого родства, так и явного сходства) и занимают обособленное положение среди хищных блестянок (Kirejtshuk & Lawrence, 1992). Некоторые виды подрода Cybocephalus Erichson, 1844 sensu stricto также известны как хищники белокрылок (Kirejtshuk, James & Heffer, 1997).
Жизненный цикл.
Жизненный цикл типичного для голометаболических насекомых типа, с метаморфозом. Для блестянок в целом характерно сравнительно быстрое личиночное развитие. Период от откладки самкой яиц до выхода из них сформировавшихся имаго, как правило, в среднем занимает около месяца. Стадия имаго - чаще долгоживущая. Для многих видов характерна поливольтинность, особенно в тропических и субтропических регионах. Зимовка обычна на дефинитивной стадии развития. Обе подвижные стадии жизненного цикла блестянок обычно активны на протяжении очень короткого периода времени, в отличие от многих других групп жесткокрылых (но исключая некоторые группы из надсемейства Cucujoidea). Именно это обстоятельство позволяет Nitidulidae захватывать и быть приуроченными к исключительно разнообразным типам субстрата, часто существуя в течение сравнительно короткого периода времени (время истекания древесного сока, короткая продолжительность периода цветения и т.д.).
Враги - хищники и паразиты, защитные приспособления, мимикрия.
Имаго большинства блестянок (особенно группы антофагов и филлофагов) часто атакуются пауками, живущими на цветах (Arachnida, Thomisidae) и перепончатокрылыми (Hymenoptera, Sphecidae). Личинки, живущие под корой, нередко поедаются птицами (Picidae). Многие виды хищных членистоногих, в особенности жуков (Coleoptera, Carabidae, наиболее часто из рода Pterostichus), а также многоножки (Chilopoda) и муравьи (Hymenoptera, Formicidae) поедают куколок блестянок - антофагов, находящиеся в почве. Личинки обитателей разлагающихся субстратов могут поедаться различными видами стафилинид (Coleoptera, Staphylinidae, в особенности видами рода Atheta) и муравьями. Взрослые жуки на лету истребляются многими видами птиц, особенно из семейств Apodidae и Hirundinidae.
Эндопаразиты блестянок также очень разнообразны и включают многие виды нематод из разных групп (особенно Tylenchida) и семейств, например, Sphaerulariidae и Tetradonematidae. Различные виды блестянок бывают заражены клещами (Acari, Trombiculidae), особенно те, которые живут под корой. Несколько видов блестянок - антофагов рода Meligethes, связанные в своем развитии с растениями семейства Brassicaceae, заражаются грегаринами и микроспоридиями, обычно Nosema meligethi I. & R. (Lipa & Hokkanen, 1992). Некоторые блестянки поражаются аскомицетами (Ascomycetes) из рода Rickia, в частности виды родов Cyllodes, Pallodes, Aethina, Platychora и Epuraea. Кроме того, блестянок могут паразитировать также Cucujomyces и Synandromyces (Audisio, 1993). Блестянки также атакуются большим числом видов паразитических перепончатокрылых семейств: Braconidae (из родов Microctonus, Blacus, Diospilus, Eubadizon, Leiophron, Microgaster), Encyrtidae (из рода Zeteticontus), Ichneumonidae (из родов Phradis, Tersilochus, Aneucles, Allophris и Stethantyx), Proctotrupidae (из рода Brachyserphus), Bethylidae (из рода Pseudisobrachium), Pteromalidae (из родов Habrocytus, Chrysolampus и Anosopteromalus) и Torymidae (из рода Totymus). Большинство атакуемых видов являются хозяйственно значимыми и поэтому исследованными видами (Carpophilus spp., Stelidota spp., Meligethes группа Meligethes aeneus и др.) (Williams et al., 1984). Та же закономерность наблюдается и среди видов рода Meligethes, атакуемых большим числом видов микроспоридий. Особенно интересными являются перепончатокрылые паразиты из семейств Encyrtidae (род Zeteticontus) и Braconidae (Microctonus nitidulus Loan), которые, по-видимому, могут быть использованы для контроля численности вредящих видов. Некоторые виды двукрылых (Diptera, Tachinidae, например, Hyalomyodes triangulifer в США) являются паразитами крупных блестянок, обычно Glischrochilus qadrisignatus (Say).
Наиболее интересные случаи мимикрии среди представителей Nitidulidae встречаются у видов из родов Soronia Erichson, 1843, Cryptarcha Shuckard, 1839, обитающих на натеках древесного сока и практически полностью сливающихся с корой, благодаря рисунку из разорванных пятен и перевязей. Виды родов Omosita Erichson, 1843 и Nitidula Fabricius, 1775 обитают на костях, трупах и шкурах животных и имеют окраску под цвет субстрата: желтый или коричневый фон тела с перевязями, пятнами или черточками, разбросанными по надкрыльям. Виды Epuraea spp. и многие другие часто остаются незаметно сидящими на трутовике или коре, за счет рыжей или темно - коричневой окраски, а представители родов Glischrochilus Reitter, 1873 и Ipidia Erichson, 1843, живущие под корой, наоборот, демонстрируют свои ярко - оранжевые или желтые пятна на надкрыльях, отпугивающие потенциальных хищников. Виды подсемейства Cybocephalinae интересны своей особенностью сворачиваться в шар при опасности.
В условиях Индокитая ряд видов из рода Cyllodes (C. bifascies, C. quadrinotatus и др.) могут собираться в скопления из десятков (или сотен) особей на стволах некоторых деревьев в тенистых местах тропического леса. Скопления занимают заплесневелые участки гладкой коры (например, на толстых фикусах). Кроме экземпляров Cyllodes, в состав этих скоплений часто входят представители сходно окрашенных и близких по размерам видов из семейств Coccinellidae и Tenebrionidae (из рода Derispia Lewis, 1894). Так, среди C. bifascies ловились экземпляры Aspidimerus matsumurae Sasaji, 1968 (Coccinellidae); среди C. quadrinotatus и C. undulatus - Aspidimerus chapaensis Hoang, 1982 и кокцинелловидные чернотелки Derispia lunata Kaszab, 1946; среди C. multimaculatus - Derispia ardoini Kaszab, 1980; C. decens - Derispia annamita Pic, 1921. Иногда в этих скоплениях встречались также сходно окрашенные Erotylidae и Endomychidae. Размер этих скоплений порой достигал 0.5-0.6 м в поперечнике, с плотностью особей до десятка на 1 кв. дециметр. Сами скопления хорошо выделялись в виде темных пятен на светло сером фоне коры. Возможно образование этих скоплений связано с половым поведением жуков (скопление - "сигнал" дальнего действия для привлечения половых партнеров), так как питание блестянок из рода Cyllodes, а также развитие их личинок происходит в других субстратах (мягкие трутовики, грибы типа вешанок, забродивший древесный сок, вытекающий из повреждений на стволах деревьев).
Литература
Кирейчук А.Г. 1981. Новый род жуков-блестянок подсем. Meligethinae (Coleoptera, Nitidulidae) из Южной Африки. Annales Historico-Naturales Musei Nationalis Hungarici. T. 73: 133-135.
Кирейчук А.Г. 1982. Систематическое положение рода Calonecrus J. Thomson и замечания по филогении семейства жуков-блестянок (Coleoptera, Nitidulidae). Энтомологическое обозрение, Т. 61, вып. 1: 117-130.
Кирейчук А.Г. 1985. Новые виды Cyllodes Erichson и Viettherchnus gen. n. (Coleoptera, Nitidulidae) фауны Вьетнама и сопредельных территорий. В кн.: Насекомые Вьетнама. М., Наука, 157-164.
Кирейчук А.Г. 1986. Анализ строения гениталий для реконструкции филогении и обоснования системы семейства блестянок (Nitidulidae, Coleoptera). Труды Всесоюзного Энтомологического общества, Т. 68: 22-28.
Кирейчук А.Г. 1986. Ревизия рода Aethina Er. (Coleoptera, Nitidulidae) фауны Ориентальной и Палеарктической областей. Труды Зоологического института АН СССР, Т.. 140: 44-82.
Кирейчук А.Г. 1986. Новые виды жуков-блестянок (Coleoptera, Nitidulidae) Австралийской области. I. Энтомологическое обозрение, Т. 65, вып. 3: 559-573.
Кирейчук А.Г. 1987. Новые виды жуков-блестянок (Coleoptera, Nitidulidae) Австралийской области. II. Энтомологическое обозрение, Т. 66, вып. 4: 773-799.
Кирейчук А.Г. 1987. Новые виды комплекса родов, близких к Cyllodes (Coleoptera, Nitidulidae) из Индокитая и сопредельных территорий. В кн.: Энтомофауна Вьетнама, М., Наука, 137-170.
Кирейчук А.Г. 1987. Новый для Индо-Малайской области род жуков-блестянок Anister Grouvelle (Coleoptera, Nitidulidae). В кн.: Энтомофауна Вьетнама, М., Наука, 170-172.
Кирейчук А.Г. 1987. Новые таксоны жуков-блестянок (Coleoptera, Nitidulidae) Восточного полушария (часть 1). Omosita nearctica sp.n., викарирующий с палеарктическим O. colon (L.). T. Труды Зоологического института АН СССР, Т.. 164: 63-94.
Кирейчук А.Г. 1987. Обзор жуков-блестянок подсем. Cryptarchinae (Coleoptera, Nitidulidae) из Индо-Малайской области. Труды Зоологического института АН СССР, Т.. 170: 62-95.
Кирейчук А.Г. 1988. Новые таксоны жуков-блестянок (Coleoptera, Nitidulidae) Восточного полушария. Часть 2. Труды Зоологического института АН СССР, Т.. 178: 62- 97.
Кирейчук А.Г. 1989. О становлении филлофагии (филлофагизации) среди жуков (Coleoptera). Труды Зоологического института АН СССР, Т.. 202: 147-182.
Кирейчук А.Г. 1989. Новые таксоны жуков-блестянок (Coleoptera, Nitidulidae) Восточного полушария (часть III). Труды Зоологического института АН СССР, Т.. 208: 64-89.
Кирейчук А.Г. 1990. Новые виды и замечания по таксономии жуков-блестянок (Coleoptera, Nitidulidae) Индокитая и сопредельных территорий. Часть 1. Труды Зоологического института АН СССР, Т. 209: 61-98.
Кирейчук А.Г. 1990. Новые таксоны жуков-блестянок (Coleoptera, Nitidulidae) Восточного полушария. Часть 4. Труды Зоологического института АН СССР, Т.. 211: 84- 103.
Кирейчук А.Г. 1990. Новые виды жуков-блестянок (Coleoptera, Nitidulidae) Австралийской области. III. Энтомологическое обозрение, Т. 69, вып. 4: 857-878.
Кирейчук А.Г. 1992. Значение антофагии в историческом развитии отряда жесткокрылых (в основном на примере семейства жуков-блестянок). Автореферат диссертации на соискание ученой степени доктора биологических наук, Ст-Петербург, Зоологический институт РАН, 1-51.
Кирейчук А.Г. 1992. Определительная таблица надсемейств Cucujoidea и Tenebrionoidea.; Определительная таблица семейств надсемейства Cucujoidea.; Сем. Блестянки - Nitidulidae.; Сем. Катеретиды - Kateretidae. В кн.: Определитель насекомых Дальнего Востока, Ст-Петербург, Наука, Т. 3, часть 2: 107-216.
Кирейчук А.Г. 1994. Система, эволюция образа жизни и филогения отряда жуков (Coleoptera). I. Энтомологическое обозрение, Т. 73, вып. 2: 266-288.
Кирейчук А.Г. 1994. Новые виды и замечания по таксономии жуков-блестянок (Coleoptera, Nitidulidae) Индокитая и сопредельных территорий. Часть 2. Труды Зоологического института АН СССР, Т.. 257: 92-127.
Кирейчук А.Г. 1995. Новые таксоны жуков-блестянок (Coleoptera, Nitidulidae) Восточного полушария. Часть 5. Труды Зоологического института АН СССР, Т. 257: 3- 50.
Кирейчук А.Г. 1996. Система, эволюция образа жизни и филогения отряда жуков (Coleoptera). II. Энтомологическое обозрение, Т. 75, вып. 1: 39-62. [перевод - 1997. System, evolution of mode of life, and phylogeny of the order Coleoptera. II. Entomological review, Vol. 76 (1): 1-20]
Кирейчук А.Г. 1998. Положение подсем. Maynipeplinae (Coleoptera, Nitidulidae) из Экваториальной Африки в системе, а также замечания по эволюции и структурным модификациям у блестянок. Энтомологическое обозрение, Т. 77, вып. 3: 540-554.
Кирейчук А.Г., Истон А.М. 1988. Ревизия рода Anthystrix Kirejtshuk и новые виды подсем. Meligethinae (Coleoptera, Nitidulidae) из Южной Африки. Труды Всесоюзного Энтомологического общества, Т. 70: 41-55.
Кирейчук А.Г., Кабаков О.Н. 1997. Замечания о жуках- блестянках (Coleoptera, Nitidulidae), собранных О.Н. Кабаковым во Вьетнаме и Лаосе. Известия Харьковского Энтомологического общества, Т. 5 (2): 13-23.
Кирейчук А.Г., Пономаренко А.Г. 1990. Ископаемые жуки семейств Peltidae и Nitidulidae. Палеонтологический журнал. N 2: 78-88 + 1 табл.
Пономаренко Ф.Г. Историческое развитие жесткокрылых насекомых. Автореферат диссерт. на соиск. степ. докт. биол. наук. Москва, Палеонт. инст. АН СССР, 1983 47 стр.
Якобсон Г.Г. 1915. Жуки России и Западной Европы. 11 часть. - Санкт-Петербург: Издание А.Ф.Девриена - 1915 - С. 865-1024.
Alfieri, A. 1924. Notes sur Anister raffrayi Grouv. et sa larva (Cole'opt.). Bull. Soc. R. Entom. Egypte, 15: 82-83.
Audisio, P. (1982): Contributo alla conoscenza dei Nitidulidae (Coleoptera) della Sierra Leone. - Recerche Biologiche in Sierra Leone. Accademia Nazionale dei Lincei, Qued. 255: 105-114.
Audisio, P. 1980. M'agyarorzsаg Allatvilаga. VIII. Kotet, Coleoptera III., 9 Fuzet: Nitidulidae. (Fauna Hungariae, Budapest, 140), 171 pp.
Audisio P.A. 1993. Coleoptera Nitidulidae - Kateretidae. Fauna d'Italia. Edizioni Calderini, Bologna, 32: 971 pp.
Audisio, P. 1994. The Meligethes of Sierra Leone (Coleoptera: Nitidulidae, Meligethinae). in Ricerche Biologiche in Sierra Leone ( parte IV ). Problemi Attuali di Scienza e di Cultura: Missioni ed esplorazioni - XIII. (Accademia Nazionale dei Linzei, 391), Roma, Quaderno, N 267: 343-356.
Audisio P. & Kirejtshuk A.G. 1983. Revision of the genera Ithyra Reitter and Neothalycra Grouvelle (Coleoptera, Nitidulidae). Revue de Zoologie africaine., Vol. 97, Fasc. 2: 365-378.
Audisio P.A. & Kirejtshuk A.G. 1987. La posizione sistematica di Xenostrongylus ovulum Fairmaire, 1875 e taxa correlati (Coleoptera, Nitidulidae). Fragm. Entomol. Vol. 20, fasc. 1: 71-75.
Audisio P. & Kirejtshuk A.G. 1995. Two new species of Meligethes (Chromogethes) from East Africa (Coleoptera, Nitidulidae). Fragmenta entomologica, Vol. 26 (2): 235- 332.
Audisio P. & Kirk-Spriggs A.H. & Kirejtshuk A.G. 1998. Meligethes of the pubescens species-group from Southern Africa (Coleoptera: Nitidulidae, Meligethinae). Entomologica scandinavica, Vol. 29 (2): 169-198.
Blazejewski F. 1968. Untersuchungen ueber die Entwicklung des Aedeagus bei dem Repsglanzkaefer Melegethes aeneus F. (Coleoptera, Nitidulidae). Polsk. Pismo Entomol., T. 38, Fasc. 2: 229-258.
Blackburn T. 1891. Further notes on Australian Coleoptera, with description of new genera and species. Part IX. - The Transactions of the Royal Society of South Australia 14: 65-153.
Boheman, C.H. 1851. Insecta Caffraria annis 1838-1845 a F.A. Wahlberg collecta. Stockholm, Holmiae 1, 2: 299-626.
Borges P.A.V. 1990. A checklist of the Coleoptera from the Azores with some systematic and biogeographic comments. -Bol. Mus. Mun. Funchal 42 (220), 87-136.
Boeving, A.G., Craighead, F.C. 1931. An illustrated synopsis of the principal larval forms of the order Coleoptera. Entomologica Americana (new series) 11: 1-351.
Boeving, A.G., Rozen, J.G. 1962. Anatomical and systematic study of the mature larvae of the Nitidulidae (Coleoptera). Entomologiske Meddelelser, 31: 265-299.
Bruch C. 1923. Coleopteros fertilizadores de "Prosopache Burmeisteri" De Bari. Physis (Revista de la Sociedad Argentina de Ciencias Naturales), 7: 82-88.
Chatterjee, S.N. 1924. Catalogue of Indian Insects. Part. 5. Nitidulidae. Government of India. Centr. Publ. Br. , 40 pp.
Connell, W.A. 1957 (1956). Nitidulidae of Delaware. Delaware Agricultural Experemental Station, Bulletin, 318, 67 pp.
Connell, W.A. 1984. Nearctic Nitidulidae - synonymy and additions since Parsons revisions. Coleopterist's Buletin, 38 (2): 160-164.
Cooper M.C. 1982. The species of the genus Pria Stephens (Coleoptera: Nitidulidae). - Zool. J. Linn. Soc. 75: 327-390.
Crowson R.А., 1955. The natural classification of the families of the Coleoptera. Nathaniel Lloyd Со, Ltd, London, 187 р.
Crowson R.A., 1975. The evolutionary history of Coleoptera as documented by fossil and comparative evidence. In: Atti Х Congresso Naz. Ital. Entomol. Firenze. 47-90.
Crowson R.А., 1981. The biology of the Coleoptera. Academic Press. London-NХ.-Toronto-Sydney-San Francisco, 802 р.
Crowson R.A. 1995. Some interesting evolutionary parallels in Coleoptera. in: J. Pakaluk & S.A. Slipinski (eds.). Biology, Phylogeny, and Classification of Coleoptera. Papers celebrating the 80th Birthday of Roy A. Crowson. Warszawa, Museum i Instytut Zoologii PAN, Vol. 1: 63-85.
De Jong, D. 1953. A "new" nitidulid beetle from Sumatra. Zoologische Mededeelingen, 32: 43-47.
Delobel A. & Tran M. 1993. Les Coleopteres des denrees alimentaires antreposees dans les regions chaudes. - Orstom/CTA, Paris, Fauna tropicale, 32, 1-425.
Dobson, R.M. 1954b. The species of Carpophilus Stephens associated with stored products. Bulletin of Entomological Research, 45: 389-402.
Dobson, R.M. 1960. Notes on taxonomy and occurence of Carpophilus Stephens (Col., Nitidulidae) associated with stored products. Entomologist's Monthly Magazine, 95: 156-158.
Dobson, R.M. 1993a. New species and subspecies of Carpophilus Stephens (Coleoptera: Nitidulidae) from the Australian region. Storkia, 2: 1-12.
Dobson, R.M. 1993b. A repraisal of Carpophilus oculatus Murray, 1864 (Coleoptera: Nitidulidae) with descriptions of two new subspecies. Storkia, 2: 26-29.
Easton A.M. 1955. The Meligethes of North Africa (Coleoptera, Nitidulidae). Me'm. Soc. Sci. nat. phys. Maroc, 22: 7-70.
Easton A.M. 1957. The Meligethes (Col., Nitidulidae) of Afganistan. Entomol. Mon. Mag. 92: 385-401.
Easton, A. M. 1957. The Meligethes of Abyssinia (Coleoptera: Nitidulidae). Trans. R. Ent Soc. London, 111 (13): 367-403.
Easton, A. M. 1959. The Meligethes of East Africa (Coleoptera: Nitidulidae). Trans. R. Ent. Soc. London, 112 (11): 263-318.
Easton, A. M. 1964. Genus Meligethes Stephens (Coleoptera: Nitidulidae). Explor. Parc. Nat. Upemba, Miss. De Witte 1946-1949, Brussels, 68: 29-59.
Endroedy-Younga S. 1968. Monographie der palaearktischen Arten der Familie Cybocephalidae (Coleoptera: Clavicornia). Acta Zool. Acad. Sci. Hung., 15, 1-2: 27-115.
Endroedy=Younga S. Systematic revision and phylogeny of some Meligethinae genera from the Ethiopian region (Coleoptera, Nitidulidae). Entomol. Germ., 1978 - Bd 4 (3/4) - S. 295-316.
Erichson, W.F. 1842. Beitrag zur Insecten-Fauna von Vandiemensland, mit besonderer Beruecksichtigung der geographischen Verbreitung der Insecten. Archiv fuer Naturgeschichte, 8 (1): 83-287, pl. 4-5.
Erichson, W.F. 1843. Versuch einer systematischen Einteilung der Nitidularien. Germar Zeitschrift fuer die Entomologie, 4: 225-361.
Erichson, W.F. 1845. Naturgeschichte der Insecten Deutschland: Coleoptera. Berlin, 3, VII + 968 pp.
Gemminger, Dr. & Harold, B. de 1868. Catalogus coleopterorum hucusque descriptorum synonymicus et systematicus. Paris-London, Monachii, Sumptu E.H. Gummi, 3: 1353 pp.
Gillogly L.R. 1962. Insects of Micronesia: Nitidulidae. - Honolulu, Bishop Mus. 16, 4, 133-188.
Grouvelle, A. 1892a. Viaggio di Leonardo Fea in Birmania e regioni vicine. 50. Nitidulides, Cucuides et Parnides. 2. Annali del Museo Civico di Storia Naturale "G. Doria", Genova, 32: 833-868.
Grouvelle, A. 1897. Clavicornes nouveaux des Indes Orientales et pays voisins (Nitidulidae). Annali del Museo Civico di Storia Naturale "G. Doria", Genova, (18) 38: 342-398.
Grouvelle, A. 1906. Contribution a l'E'tude des Colйopteres de Madagascar, Nitidulidae, Colydiidae, Cucujidae, Monotomidae, Cryptophagidae, Mycetophagidae, Dryopidae, Heteroceridae. Annales de la Socie'te' Entomogique de France, 75: 67-93.
Grouvelle, A. 1908a. Cole'opte'res de la region Indienne. Rhizodidae, Trogositidae, Nitidulidae, Colydiidae, Cucujidae. (1ere mйmoire). Annales de la Socie'te' Entomologique de France, 77: 315-495, pls.
Grouvelle, A. 1908/1909. Cole'opte'res clavicornes de l'Afrique australe et orientale. Revue d'Entomologie, 27:127-206.
Grouvelle, A. 1913. Byturidae, Nitidulidae. In: W. Junk (ed.). Coleopterorum Catalogus. Berlin, 15, 56: 1-223.
Grouvelle, A. 1913. Insects of the Percy Sladen Trust expedition 1905. Coleoptera: Nitidulidae, Heteroceridae. Transactions of the Linnean Society, Ser. 2, Zoology, 16: 93-166.
Grouvelle, A. 1917. Nitidulidae des оles Philippines recolte's par C.F. Baker. 2. Philippines Journal of Sciences, D 12: 329-344.
Hayashi N. 1978. A contribution to the knowledge of the larvae of Nitidulidae occuring in Japan (Coleoptera: Cucujoidea). Insecta Matsum., N.S., 14: 1-97.
Hayashi N. Larvae. In: Morimoto, K. & Hayasi N. (eds.) - The Coleoptera of Japan in color. - Osaka: Hoikusha publishing Co., ltd. - 1986 - Vol. 1 - P. 202-218.
Heller, K.M. 1916. Die Kaefer von Neu-Caledonien und den benachtarten Inselgruppen. Nova Caledonia. Zool. 2, L. 3: 229-364.
Hinton, H.E. 1945. A monograph of the beetles associated with stored products. Brit Mus. Nat. Hist. London, 1, VIII + 1-443 pp.
Hlavac Т.F., 1972. The prothorax of Coleoptera: origin, major features of variation. Psyche, 79: 123-149.
Jelinek, J. 1980. Zur Taxonomie der asiatischen Nitidulidae und Cerylonidae (Coleoptera). Reichenbachia, 18, 14: 93-102.
Jelinek J., 1981. Review of the genus Anister Coleoptera, Nitidulidae). Acta entom. bohemoslov., 78: 183-188.
Jelinek, J. 1978. Ergebnisse der Bhutan-Expedition 1972 des Naturhistorischen Museums in Basel. Coleoptera: Fam. Nitidulidae. Entomologica Basiliensia, 3: 171-218.
Jelinek J. 1979. Revision of the genus Epuraea Er. from Africa with remarks to related genera (Col., Nitidulidae). -Acta Entomol. Mus. Nat. Pragae, Vol. 39 (1977), 345-397.
Jelinek, J. 1992. Nitidulidae (Coleoptera) associated with flowers of oil palm, Elaeis guineensis (Arecales, Arecacaea), in Rwanda. Acta entom. bohemosl., 89: 409-428.
Kirejtshuk A.G. 1980. New species of beetles of the subfam. Meligethinae (Coleoptera, Nitidulidae) from the Ethiopian Region. Revue de Zoologie africaine, Vol. 94, fasc. 2: 249-294.
Kirejtshuk A.G. 1981. Preliminary revision of the Cryptarchinae genera of the Afrotropical region, with description of a new genus, a new subgenus and some new species (Coleoptera, Nitidulidae). Revue de Zoologie africaine, Vol. 95, fasc. 4: 766-805.
Kirejtshuk A.G. 1984. New taxa of Nitidulidae (Coleoptera) from the Indo-Malayan fauna. Annales Historico-Naturales Musei Nationalis Hungarici, T. 76: 169-195.
Kirejtshuk A.G. 1986. On polyphyly of the Carpophilinae with description of a new subfamily, Cillaeinae (Coleoptera, Nitidulidae). Coleopterist`s Bulletin, Vol. 40, N 3: 217-221.
Kirejtshuk A.G. 1991. Evolution of mode of life as the basis for division of the beetles into groups of high taxonomic rank. In: M. Zunino, X. Belles, M. Blas (Eds.). Advances in Coleopterology, AEC, Barcelona: 249-262.
Kirejtshuk A.G. 1994. Revision of the genus Neopallodes Reitter 1884 (Coleoptera Nitidulidae) from the Palaearctic and Indo-Malayan regions. Tropical Zoology, Vol. 7, 2: 225-253.
Kirejtshuk A.G. 1996. Some results of study on the Nitidulidae from Namibia and adjacent territories. Part 1 (Coleoptera, Cucujoidea, Nitidulidae). Mitteilungen aus dem zoologischen Museum in Berlin, Bd. 76 (1996), Hf. 1: 51-82.
Kirejtshuk A.G., A.H. Kirk-Spriggs 1996. Viettherchnus Kirejtshuk, 1985 and Ceramphosia gen. n. from the Indo-Malayan region (Coleoptera, Nitidulidae). Zoosystematica Rossica. Vol. 4 (1): 131-138.
Kirejtshuk A.G., J. Pakaluk 1996. Notes on the Nearctic Epuraeinae (Coleoptera, Nitidulidae). Zoosystematica Rossica, Vol. 4 (1): 139-152.
Kirejtshuk A.G., James D.G., Heffer R. 1997. Description and biology of a new species of Cybocephalus Erichson (Coleoptera: Nitidulidea), a predator of Australian citrus whitefly. Journal of the Australian Entomological Society, Vol. 36: 81-86.
Kirejtshuk A.G. 1997. On the evolution of anthophilous Nitidulidae (Coleoptera) in tropical and subtropical regions. Bonner Zoologische Beitrage, Bd. 47: 111-134.
Kirejtshuk A.G. 1998/1999. Nitidulidae (Coleoptera) of the Himalayas and Northern Indochina. Part 1: subfamily Epuraeinae. Koeningstein, Koeltz Scientific Books (Theses Zoologicae, Vol. 28): 1-489.
Kirejtshuk A.G. 2000. On origin and early evolution of the supefamily Cucujoidea (Coleoptera, Polyphaga) Comments on the family Helotidae. The Kharkov Entomological Society Gazette, Vol. 8 (1): 8-38.
Kirejtshuk A.G. & Audisio P. 1995. Preliminary revision of South African Meligethes, subg. Lariopsis Kirejtshuk (Coleoptera, Nitidulidae, Meligethinae). Fragmenta entomologica, 1995, Vol. 27 (1): 191-154.
Kirejtshuk A.G. & Lawrence J.F. 1990. Revision of Australian genus Idaethina Reitter (Coleoptera: Nitidulidae). Journal of the Australian entomological Society, Vol. 29: 1-9.
Kirejtshuk A.G. & J.F. Lawrence 1992. Cychramptodini, new tribe of the Nitidulidae (Coleoptera) from Australian region. Journal of the Australian entomological Society, Vol. 31: 29-46.
Kirejtshuk A.G. & J.F. Lawrence 1992. Review of the Thalycrodes-complex of genera (Coleoptera, Nitidulidae), endemic to the Australian region. Journal of the Australian entomological Society, Vol. 31: 119- 142.
Kirejtshuk A.G. & Lawrence J.F. 1999. Aethina complex Notes on the Aethina complex (Coleoptera: Nitidulidae: Nitidulinae) with a review of the Aethina (Cleidorura) subgen.n. and Aethina (Idaethina) Gemminger & Harold. Annales Zoologii, 49: 000-000.
Kirejtshuk A.G., Leschen R.A.B. 1998. Review the Thalycra complex (Coleoptera: Nitidulidae) with three new genera and notes on mycophagy. Annales Zoologii, 48 (3/4): 253-273.
Kurosawa Y., S.Hisamatsu and H.Sasaji. 1985. Coloured illustrations of the Coleoptera of Japan. Hoikusha Publishing Co., Ltd, Osaka. III: 1-500.
Lacordaire, T. 1854. Histoire naturelle des Insectes. Genera des Cole'opte'res ou expose' me'thodique et critique de tous les genres propose's jusq'ici dans cet ordre d'Insectes. - Libraire Encyclope'dique de Roret 2: pp. 1-548. Paris (Roret)
Lawrence J.F., 1991. Coleoptera Nitidulidae (Cucujoidea) (including Brachypteridae, Cateretidae, Cybocephalidae, Smicripidae). pp. 456-460, in: Stehr. F.K. (ed.), Immature Insects. Kendall-Hunt Publ. Comp., Dubuque (Iowa), vol. 2 (1991). XVI + 975 pp.
Lawrence J.F. & Kirejtshuk A.G. 1999. Nitidulidae, major part.; Maynipeplinae, Nitidulidae.; Calonecrinae, Nitidulidae.; Cryptarchinae, Nitidulidae.; Cybocephalinae, Nitidulidae.; Helotidae.; Kateretidae.; Smicripidae.; Rhizophaginae, Monotomidae. In: CD by J.F. Lawrence, A.M. Gastings, M.J. Dallwittz, T.A. Paine & E.J. Zurcher "Beetles of the world: descriptions, identification and information for retrieval for families and subfamilies". DELTA, CSIRO, Entomology.
Lechanteur F. (1955): Contribution a l'etude des Brachypeplus d'Afrique (Col., Nitidulidae). - Revue de Zoologie et Botonique africaine, 52: 127-143.
Lesne P. Sur un Nitidulide myce'tophage nouveau et sur quelques caracte`res de la familie (Coleoptera). Rev. Franc. Entomol.,1938, Vol. 5, N 3, p. 158-168.
Lundie, A.E. 1940. The small hive beetles Aethina tumida. Union South Africa Governm. Printer, Pretoria, 1-30.
MacLeay, W.J. 1873. Notes on a collection of insects from Gayndah. Transactions of the Entomological Sociely of New South Wales (1871), 2: 79-205.
von Mannerheim, C.G. 1843. Beitrag zur Kaeferfauna der Aleutischen Inseln, der Inseln Sitkha und Neu-Californiens. Bulletin de la Socie'te' Impe'riale des Naturellistes de Moscou, 16 (2): 175-314.
von Mannerheim, C.G. 1852. Zweiter Nachtrag zur Kaefer-Fauna der nord-americanischen La"nder des russischen Reiches. Bulletin de la Socie'te' Impe'riale des Naturellistes de Moscou, 25 (1): 283-387.
Motschulsky, V. 1858. E'tudes Entomologiques. Helsingfors, 7: 192 pp. + Taf. 2.
Motschulsky, V. 1860. E'tudes Entomologiques. Helsingfors, 9: 42 pp.
Nunberg, M. 1976. Klucze do oznaczania owado'w polsli. Polskie Towarzystwo Entomologiczne, Warszaw: Pan'stwowe Wydawnictwo Naukowe, 19, 65, 92 pp.
Parsons C.T. 1943. A revision of Nearctic Nitidulidae (Coleoptera). Bull. Mus. Compar. Zool., 92 (3): 121-278 + 13 pl.
Paulian, A. & Villiers A. (1940): Les Cole'opte`res des Lobelias des Montagnes du Cameroun. - Revue franC,aise d'Entomologie, 7 (2): 72-83.
Rebmann, O. 1956a. Revision der Meligethes, Subgenus Odontogethes (Col., Nitid.). Entomologisch. Bl., 52: 42-48.
Rebmann, O. 1956b. Revision der Meligethes (Col., Nitid.). Die Palaearktischen Arten der Gruppe des M. atratus. Entomologische Blaetter fuer Biologie und Systematik der Kaefer (Berlin), 52: 124-135.
Reitter, E. 1873b. Systematische Einteilung der Nitidularien. Verhandlungen des naturforschenden Vereines in Brunn, 12, 1: 5-194.
Reitter, E. 1975. Die europaeischen Nitidularien mit kurzer Charakteristik der Gattungen und Bemerkungen ueber schwierige Arten verzeichnet. Deutsche Entomologische Zeitung, 19 (3): 1-30, 87-88.
Reitter E. 1884a. Die Nitiduliden Japans. Wiener Entomologische Zeitung, 3, 1-2: 257-272, 299-302.
Reitter E. 1911. Fauna Germanica. Die Kaefer des Deutschen Reiches. III. K.G.Lutz' Verlag, Stuttgart, 436 SS. + 48 Taf.
Reitter, E. 1919. Nitidulidae und Byturidae. Bestimmungs-Tabelle der europaeischen Coleopteren, 86 (Verhandlungen des naturforschenden Vereines in Bruenn, 56): 1-104.
Rozen, J.G. 1963. Preliminary systematic study of the pupae of the Nitidulidae (Coleoptera). American Museum, Novitates, 2124: 1-13.
Scholtz, M. F. R. 1932. Eine neue Meligethes aus Suedeuropa und Bemerkungen zu einigen arten. Ent. Bl. , 28: 97-100.
Schulze, F.E., Kukenthak, W., Heider, K., 1931. Nomenclator animalium generum et subgenerum. Berlin, 3: 1779-2248.
Sharp, D. 1889-1891. Fam. Nitidulidae. in: Biologia Centrali-Americana. Insecta. Coleoptera. 2, 1: 265-388, 8-12 tabs.
Sharp, D. & Scott, H. 1908. Fam. Nitidulidae. in: Sharp, D., Scott, H. 1908/1909. Fauna Hawaiiensis. Coleoptera. Cambridge, III, 3, 5: 435-508 + 13-16 pls.
Sharp D. & Muir F. 1912. The comparative anatomy of the male genital tube in Coleoptera. Transactions R. Entomological Soc. London, Vol. 3: 477-649 + 78 pls.
Shuckard, W.E. 1839. Elements of British Entomology, containing a General Introduction to the Science, a Systematic Description of all the Genera, and a List of all the Species of British Insects, with a History oftheir Transformation, Habits, Economy... Part 1. Hyppolyte BailliКre, London, 240 pp.
Shuckard, W.E. 1840. British Coleoptera delineated by W.J. Spry consisting of figures of all the Genera of British Beetles. Crosa Ed., London, 83 pp., 94 pls.
Silvestri F. 1910. Metamorphosi del Cybocephalus rufifrons Reitter e notizie sui suoi costumi. Boll. Labor. Zool. Gener. Agrar., 4: 221-227.
Sjoeberg, O. 1939. Beitrag zur Kenntnis der Gattung Epuraea Er. (Col., Nitidulidae). Bestimmungstabelle der palaearktische Arten. Entomol. Tidskr., 60: 108-126.
Spornraft K. 1967. 50. Familie: Nitidulidae. In: H. Freude, K.W. Harde & G.A. Lohse (eds.). Die Kafer Mitteleuropas,7: 20 - 77. Krefeld: Goecke & Evers Verlag.
Spornraft K. 1992. Nitidulidae, Kateretidae. In: G.A. Lohse & W.H. Lucht, Die Kaefer Mitteleuropas, 2, Supplement band mit katalogteil. Krefeld: Goecke & Evers Verlag, 13: 90-11.
Spornraft K. & A.G. Kirejtshuk 1993. Ueber alte und neue sudafrikanische Meligethes-Arten (Coleoptera, Nitidulidae). Mitteilungen Munch. Entomologische Gesselschaft Bd. 83: 47-75.
Tian M. 1995. Description of one new species of genus Cybocephalus from China (Coleoptera: Cybocephalidae. J. South China Agricultural University, 16 (2): 42-43.
Tian M.-Y. 1996. A new species of the genus Cybocephalus from Dinghushan Natural Reserve, Guangdong, China (Coleoptera: Cybocephalidae). Acta Zootaxonomica Sinica, 21 (1): 92-94. (in Chinese with English Abstract)
Tian M.-Y. 2000. Two new species of Cybocephalidae (Coleoptera) from Northwest China. Entomologia Sinica, 7 (2): 127-131.
Tian M.-Y. 2001. Two new species of Cybocephalidae from Gansu Province of China (Coleoptera: Cybocephalidae). Acta Zootaxonomica Sinica, 26 (1): 43-45. (in Chinese with English Abstract)
Tian M.-Y. & Pang H. 1994. A new species of Cybocephalus from Taiwan (Coleoptera: Cybocephalidae). Chinese J. of Entomologiy, 14: 401-403.
Tian M.-Y. & Pang H. 1997. Notes on the genus Cybocephalus from Hainan Island, China (Coleoptera: Cybocephalidae. J. South China Agricultural University, 18 (1): 34-38.
Tian M.-Y. & Ramani S. 2003. Description of a new species of Cybocephalus Erichson (Coleoptera: Cybocephalidae) from India feeding on the spiralling whitefly, with notes on its biology. Entomon, 28 (1): 21-25.
Tian M.-Y. & Yu G.Y. 1994. Two new species of of the genus Cybocephalus from China (Coleoptera: Cybocephalidae). Entomologica Sinica, 1 (3): 209-213.
Wang D.W., Huang L.L. & Zhang Q.B. 1984. Studies on the biological characteristics of Cybocephalus nipponicus Endroedy-Younga (Coleoptera: Cybocephalidae), an important predator of the arrowhead scale. Acta Phytophylastica Sinica, 11: 23-28.
Williams R.N., Fickle D.S., Kehat M., Blumberg D., Klein M.G. 1983. Bibliography of the genus Carpophilus Stephens Coleoptera: Nitidulidae. Ohio State Univ., Wooster, Research Circular 278: 1-95.
Wollaston, T.V. 1854. Insecta Maderensia: being an account of the insects of the islands of the Madeiran group. London, 634 pp.
Wollaston, T.V. 1865. Coleoptera atlantidum, being an enumeration of the coleopterous insects of the Madeiras, Salvages, and Canaries. J.V. Voorst, London, 1-140.
Wollaston, T.V. 1867. Coleoptera Hesperidum, being an enumeration of the coleopterous insects of the Cape Verde Archipelago. London, 1-285.
Yu G. 1994. Three new species of Cybocephalidae (Coleoptera) from China. Zoological Research, 15 (3): 13-17. (In Chinese with English Abstract)
Yu G. 1995a. Two new species of Cybocephalidae from Guangdong, China (Coleoptera). Entomotaxonimia, 17 (1): 31-34.
Yu G. 1995b. Two new species of Cybocephalidae (Coleoptera) from China. Entomotaxonimia, 17 (4): 277-280.
Yu G. 1997. Coleoptera: Cybocephalidae. In: "Insects of the Three Gorge Reservoir area of Yangtze River". Chongqin Pub. House, 665-668.
Yu G. & Tian M.Y. 1995. Notes on the genus Cybocephalus from China (Coleoptera: Cybocephalidae). Entomologica Sinica, 2 (1): 35-38.
Последнее обновление
Декабрь 2004 г.