
|

|




|
|
|
|
|
|
|
|
Саратовский государственный университет им. Н.Г. Чернышевского
Саратовское отделение Русского энтомологического общества
E-mail: sazh@list.ru
Семейство Heteroceridae MacLeay, 1825
Число видов в мире и России.
Пилоусы (Heteroceridae MacLeay, 1825) - небольшое всесветно распространённое (кроме Антарктиды) семейство жесткокрылых. Русское название "пилоусы" гетероцериды получили из-за явной пильчатости булавы усиков. Мировая фауна семейства не ревизована, однако, известно более 330 видов. Виды в основном группируются, по разным классификациям, либо в два подсемейства: Heterocerinae и Elythomerinae, с выделением 15 родов [Vanin et al., 2005]; либо в одно подсемейство Heterocerinae с 5 родами [Hallan, 2008]. Наиболее богата фауна тропической и субтропической климатических зон.
В Палеарктике отмечено 93 вида из 3 родов: Augyles (60 видов), Heterocerus (31 вид), Micilus (2 вида) [Mascagni, 2006].
На территории России на данное время отмечено 19 видов семейства из двух родов (Augyles, Heterocerus) [Сажнев, 2013], из них 14 видов - в Европейской части страны [Зайцев, 1916; Кирейчук, 2001; Сажнев, 2013; Zaitsev, 1908; Mascagni, 2006]. Вероятно нахождение новых для фауны России видов из родов Augyles, Heterocerus, Micilus.
Морфология.
Имаго. Длина от 1 до 8 мм. Тело вытянутое, умеренно уплощенное, в основном густо пунктировано, покрыто прилегающими и/или торчащими шелковистыми волосками, по бокам обычно имеется ряд из более длинных волосков (рис. 1). Тело самок более компактно и короче, чем у самцов. Окраска, как правило, черная, коричневая или бурая, надкрылья обычно с рисунком из различного вида пятен.

Рис.1. Габитус имаго Heterocerus fossor (фотография К.В. Макарова) |
Голова прогнатная с умеренно крупными развитыми сложными округлыми глазами по бокам и сильно выступающими мандибулами. Фронтоклипеальный шов очень разнообразен. Наличник самцов часто несет небольшие рожки (рис. 2). Верхняя губа крупная, с плотными щетинками по переднему краю, на вершине часто вырезанная. Мандибулы изогнутые, заострённые, с несколькими зубчиками на режущем крае, часто стирающимися, наружный край несет ряд щетинок. У ряда видов мандибулы самцов увеличены (Heterocerus parallelus, H. gnatho и др.), что является выражением полового диморфизма (рис. 3). Челюстные щупики четырёхсегментные, тонкие, умеренной длины. Субментум большой, продолговатый, глубоковыемчатый на центральном крае; язычок кожистый, двухлопастный, лишён параглосс; губные щупики тонкие, трёхсегментные [Егоров, 1989; Кирейчук, 2001; Vanin et al., 2005].
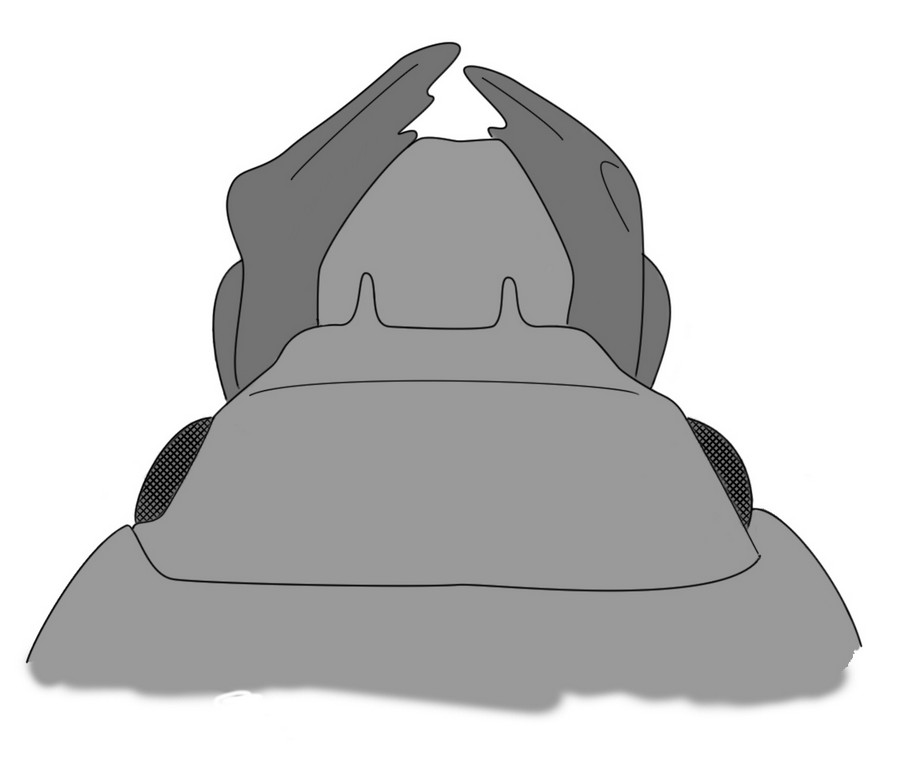
Рис. 2. Голова самца Heterocerus fenestratus с клипеальными рожками и увеличенными мандибулами (по Литовкин, Сажнев, Клёмин, 2013 (в печати)). |

Рис. 3. Половой диморфизм Heterocerus parallelus: слева - самец, справа - самка (по Литовкин, Сажнев, Клёмин, 2013 (в печати)). |
Усики 9- или 11-члениковые, сильно измененные, с третьего или пятого сегмента слабопильчатые (рис. 4), образуют удлинённую свободную 6- или 7-члениковую булаву, расположены между глазами и основанием мандибул.

Рис. 4. Усик Efflagitatus freudei (по Vanin et al., 2005). |
Переднеспинка шире головы, поперечная, по бокам округлая, почти трапециевидная до почти эллиптической, максимально широкая у основания или в задней трети. Опушенная, с плотной боковой бахромой из длинных волосков. С закруглёнными или выступающими задними углами, со слабо заострённым боковым краем, поверхность пунктирована. Основание переднеспинки прямое или в середине она чуть оттянута назад. Вдоль основания переднеспинки иногда имеется выраженная складка, доходящая до её задних углов.
Переднегрудь вытянута вперёд, её передний край слабовыемчатый, прикрывает нижнюю часть головы. Передние тазики расположены у основания переднегруди, разделены узким выпуклым продолговатым килем, уплощающимися к переднему краю переднегруди. Тазиковые впадины широко открыты в задней их части. Трохантин передних лапок явный; передние тазики широкие, поперечные, узко разделённые.
Среднегрудь короткая, заднегрудь длинная. Щиток маленьких размеров, треугольной формы, по направлению к вершине заострён. Средние тазики шаровидные, разделены шире, чем передние. Задние тазики узкие, поперечные, почти соприкасающиеся, имеют бедренные покрышки. Трохантеры большие, треугольной формы. Бёдра слабо уплощённые [Егоров, 1989; Кирейчук, 2001; Vanin et al., 2005].
Формула лапок 4-4-4, их сегменты нитевидные, с длинными редкими волосками на нижней стороне, тарсомер 4 умеренно развит и почти такой же длинный, как тарсомер 1. Коготки длинные, тонкие. Передние ноги копательного типа. Голени передних и задних ног расширенные, уплощённые и вооружены рядом мощных шипов; задняя пара несёт более тонкие шипы.
Надкрылья неукороченные, выпуклые, с совместно округлёнными вершинами. Бороздки слабо вдавленные, неотчётливые. Поверхность промежутков мелко пунктированная, нередко пунктировка двойная. Эпиплевральное поле занимает весь боковой край надкрылий, к вершине сужается. На надкрыльях почти всегда имеется рисунок из пятен, светлых на темном фоне, или темных на светлом. Строение рисунка обычно закономерно, однако весьма изменчиво, пятна могут сливаться в перевязи и наоборот (рис. 5).

Рис. 5. Схема элементов рисунка надкрылий рода Heterocerus: слева направо - H. fenestratus; H. flexuosus; H. marginatus. Обозначения, принятые на рисунке: S-пв - S-образная перевязь; N-спв - N-образная срединная перевязь; U-пп - U-образное плечевое пятно; кк - краевой кант; пвп - предвершинное пятно; ппп - предвершинная пара пятен; пшп - пришовное пятно; пщп - прищитковая полоса; спп - срединная пара пятен (по Литовкин, Сажнев, Клёмин, 2013 (в печати)). |
Брюшко с пятью видимыми стернитами, швы менее отчётливые в середине между первым и вторым видимыми стернитами, иногда эти стерниты сливаются. Первый стернит несёт боковые изогнутые кили (для стридуляции) (рис. 6). Бедренные линии различают полные (линия имеет форму половины окружности - род Augyles) и неполные (линия имеет вид лишь четверти окружности - род Heterocerus).

Рис. 6. Строение бедренных линий пилоусов двух родов: 1 - Augyles hispidulus; 2 - Heterocerus fenestratus (по Литовкин, Сажнев, Клёмин, 2013 (в печати)). |
Личинка. Личинки удлиненные и сильновыпуклые, с более широкими грудными, несколько сдавленными по бокам брюшными сегментами (рис. 7). Длина тела 2 - 10 мм. Личинки камподеевидного типа, подвижные. Спинная поверхности головы и тергиты темно-коричневые, в торчащих коротких и длинных щетинках, голова, грудь и тергиты I-IV с более густо расположенными короткими хетами.
Голова прогнатная, с очень короткими ротовыми частями
и усиками. Усики трёхчлениковые. Мандибулы симметричны, на вершине раздвоены. Максиллярные
щупики трёхсегментные, нижнегубные - двух.
Переднегрудь с развитой переднеспинкой, плевриты гладкие, в виде мембран.
Средне- и заднегрудь уже переднеспинки. Ноги состоят из 5 сегментов, длина их
слегка увеличивается от первой к третьей паре. Передние конечности роющего
типа.

Рис. 7. Личинка Efflagitatus freudei (по Vanin et al., 2005). |
Брюшко продолговатое, состоит из 10 сегментов. 9-й брюшной сегмент широко закруглен на вершине, без крышечки, без урогомф; брюшко без жабр [Кирейчук, 2001; Vanin et al., 2005].
Личинки и куколки многих видов еще не описаны.
Куколка. Длина 3 - 8 мм. Куколки продолговато-овальные, беловатые, свободные (рис. 8). Густо опушены, с короткими и длинными щетинками. Голова сверху не видна. Переднеспинка поперечная, с сильно выпуклым передним краем. Птеротеции опушенные, тесно прижаты к телу, кзади расширяются. Ноги голые. Брюшные сегменты I-VIII с поперечными рядами щетинок.

Рис. 8. Куколка Efflagitatus freudei (по Vanin et al., 2005). |
Куколки развиваются в грязевом коконе. Располагаются в колыбельке обычно на спине, на конце брюшка часто присутствуют остатки личиночного экзувия.
Экология и биология
На всех стадиях развития жуки приурочены к берегам водоёмов различных типов и с разной степенью солёности, обычно заселяют узкую полосу вдоль уреза воды. Являются характерными интерстациальными обитателями краевых структур «вода-суша». Предпочитают незаросшие или малозаросшие участки берега с умеренным увлажнением. Заселяют дисперсные, часто заиленные, виды грунта: песчаные и/или глинистые. Как правило, зона уреза обитаема жуками не более, чем на 1 м от уреза воды. На крупных реках зона может распространяться и далее.
Имаго и личинки сооружают в субстрате разветвлённые сети тоннелей, которые различаются по строению и служат разным целям: питанию, откладке яиц, окукливанию, зимовке (рис. 9). Яйца откладываются группами. Часто образуют колонии, нередко разные виды обитают синтопно. Колонии весьма мобильны, т.к. тесно связаны с ритмами колебания воды, и, в зависимости от степени увлажнения, меняют свое положение относительно уреза. При затоплении тоннелей имаго покидают их и быстро передвигаются по поверхности субстрата, могут совершать небольшие перелёты. При осушении временных водоёмов или длительном затоплении береговой линии жуки образуют временные колонии, где не размножаются, или же меняют место обитания.

Рис. 9. Различные типы тоннелей гетероцерид (по Kaufmann, Stansly, 1979) |
Есть также сведения о встречах пилоусов под водой на значительном удалении от берега и в бентосе. Личинки и имаго - альго- и фитодетритофаги, но имеется мнение о питании также зоопланктоном [Кирейчук, 2001; Прокин, 2008; Pacheco, 1978; Van Strien, 1980; Vanin et al., 2005]. Имаго многих видов летят на искусственный свет.
Согласно классификации жизненных форм (на основе наиболее изученного семейства жужелиц (Carabidae)) гетероцерид можно отнести, как по морфологии, так и по образу жизни, к группе роющих геобионтов - специализированных к рытью насекомых. Учитывая подобную биономию, а также зависимость от субстрата (пилоусам необходима влажность от 30 до 70% и определенная структура грунта, эти условия выполняются именно в зоне уреза воды) можно считать данное семейство (совместно с рядом сходных по образу жизни жесткокрылых) «маркерами» маргинальных участков экотонов «вода - суша».
В целом экология семейства изучена поверхностно. Учитывая сборы из ряда регионов России и зарубежья, можно предположить, что большинство видов семейства предпочитают песчаные и супесчаные почвы, причем выбор водных объектов в большей степени зависит от географического положения, в аридных зонах жуки зачастую заселяют солончаки и водоемы с умеренной соленостью, хотя исключительных галофилов в этом семействе нет. Морские побережья и солончаки заселяют Heterocerus fenestratus, Heterocerus parallelus и другие. На песках обитают Augyles hispidulus, Heterocerus marginatus и прочие.
Большинство видов семейства, отмеченных для России, обладают евро-сибирскими типами ареалов. Наиболее распространенным и обычным в России является Heterocerus fenestratus - эвритопный вид, который заселяет берега различных водных объектов, включая и антропогенные ландшафты. Встречается на огромной территории - имеет транспалеарктический полизональный ареал.
Роль в природе
Пилоусы, как детритофаги, являются важным звеном в круговороте веществ в живой природе, усиливая доступность органики для бактерий и иных организмов. Определенно участвуют в переносе вещества и энергии в зоне перехода двух сред - воды и суши. Гетероцериды служат важным элементом питания для воробьиных [Schmidt et al., 2004], амфибий [Turner, 1959] и других животных, по-видимому, они также играют определенную роль в закапывании и прорастании семян на песчаных почвах [Bernhardt, 1995]. На представителях семейства паразитируют клещи группы Hydrachnidiae, а также грибы отряда Laboulbeniales, в частности Botryandromyces ornatus.
Методы сбора
Пилоусы покидают свои норы после захода солнца, в это время наиболее эффективным способом сбора является привлечение их на источник искусственного света (ДРЛ и УФ-лампы), расположенный недалеко от берега водного объекта. Особенно продуктивным является сбор на светоловушки с накопителем (контейнерные). Жуки хорошо летают, а при попутном ветре нередко ловятся и вдали от водоемов.
В светлое время суток имаго и личинок можно добыть при помощи общепринятых методик, основанных на отпугивании беспозвоночных посредством создания на исследуемом участке повышенной влажности - выплёскивания и вытаптывания.
Неплохие результаты дает просеивание субстрата через энтомологическое сито - способ, связанный с активным механическим воздействием на субстрат, в результате чего происходит его разделение на фракции. Живые объекты в дальнейшем выбираются вручную. Просеиванию предшествует изъятие части субстрата вдоль береговой линии при помощи скребка.
Учитывая большую изменчивость рисунка у некоторых видов, проводить определение рекомендуется по серийному материалу. Многие виды хорошо различаются по строению тегмена, входящего в состав копулятивного аппарата самца.
Фотографии многих видов гетероцерид можно посмотреть в
составляемом мной атласе, а также в атласе семейства Heteroceridae на этом сайте.
Литература
Егоров А.Б., 1989. Сем. Heteroceridae - Пилоусы // Определитель насекомых Дальнего Востока СССР. Т. 3. Жесткокрылые, или жуки. Ч. 1. Л. С. 451-453.
Зайцев Ф.А., 1916. Заметки о некоторых представителях семейства Heteroceridae (Coleoptera) // Русское энтомологическое обозрение. Т. 15. No. 4. С. 569-571.
Кирейчук А.Г., 2001. Семейство Heteroceridae (Пилоусы) // С.Я. Цалолихин (ред.): Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые (ручейники, чешуекрылые, жесткокрылые, сетчатокрылые, большекрылые, перепончатокрылые). СПб.: Наука. С. 341-348.
Прокин А.А., 2008. Водные жесткокрылые (Coleoptera) малых рек Европейской части России: разнообразие, биоценотическая и индикационная роль // Экосистемы малых рек: биоразнообразие, экология, охрана. Лекции и материалы докладов Всероссийской школы-конференции. Института биологии внутренних вод им. И.Д. Папанина. 18-21 ноября 2008 г. С. 38-53.
Сажнев А.С., 2013. Каталог видов семейства Heteroceridae (Пилоусы) фауны России // http://www.zin.ru/ANIMALIA/COLEOPTERA/RUS/heter_ru.htm
Bernhardt K., 1995. Seed burial by soil burrowing beetles // Nordic Journal of Botany 15: 257-260.
Hallan J., 2008. Biology catalog. World Wide Web electronic publication - http://entowww.tamu.edu/research/collection/hallan
Kaufmann T., Stansly P.A., 1979. Bionomics of Neoheterocerus pallidus Say (Coleoptera:а Heteroceridae) in Oklahoma. J. Kansas Entomol. Soc. 52: 565-577.
Mascagni A., 2006. Family Heteroceridae MacLeay, 1825 // I. Löbl, A. Smetana (eds.): Catalogue of Palaearctic Coleoptera. Vol. 3. Scarabaeoidea, Scirtoidea, Dascilloidea, Buprestoidea, Byrroidea. Stenstrup: Appolo Books. P. 446-449.
Pacheco F., 1978. A catalog of the Coleoptera of America north of Mexico. Family: Heteroceridae // United States Department of Agriculture. Agriculture handbook. Washington: United States Department of Agriculture. Vol. 529-547. 8 pp.
Schmidt M., Lefebvre G., Poulin B., Tscharntke T., 2004. Reed cutting affects arthropod communities, potentially reducing food for passerine birds. Biological Conservation 121: 157-166.
Turner F.B., 1959. An analysis of the feeding habits of Rana p. pretiosa in Yellowstone Park, Wyoming. The American Midland Naturalist 61: 404-413.
Van Strien A.J., 1980. De Nederlandse soorten van de keverfamilie Heteroceridae // Zoologische Bijdragen. Vol. 27. P. 9-42.
Vanin S.A., Costa C., Ide S., Beutel R.G., 2005. Heteroceridae MacLeay, 1825 // Handbook of Zoology. Vol. 4. Arthropoda: Insecta. Pt. 38. Coleoptera, Beetles. Vol. 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Berlin, New York: Walter de Gruyter. P. 518-521.
Zaitzev, Ph.A., 1908. Catalogue de Coleopteres agutigues des familles de Dryopidae, Georyssidae, Cyathoceridae, Heteroceridae et Hydrophilidae // Horae Societitas Entomologicae Rossicae. Vol. 38, No. 4. P. 283-420.